Download
1 / 1
10 likes | 191 Views
Contribution of inactivated influenza virus (IIV) and live, attenuated influenza virus (LAIV) vaccine-induced immunity toward limiting influenza:GAS super-infections. P-569.

E N D
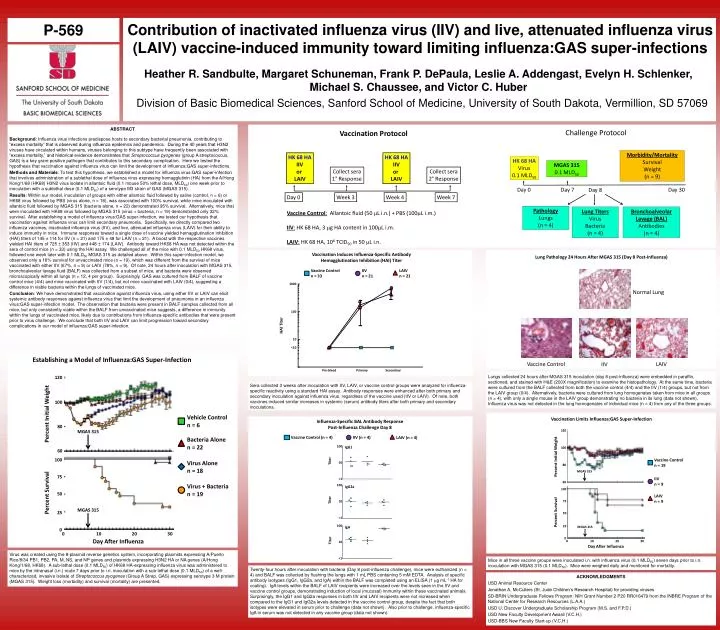
Contribution of inactivated influenza virus (IIV) and live, attenuated influenza virus (LAIV) vaccine-induced immunity toward limiting influenza:GAS super-infections P-569 Heather R. Sandbulte, Margaret Schuneman, Frank P. DePaula, Leslie A. Addengast, Evelyn H. Schlenker, Michael S. Chaussee, and Victor C. Huber Division of Basic Biomedical Sciences, Sanford School of Medicine, University of South Dakota, Vermillion, SD 57069 ABSTRACT Background: Influenza virus infections predispose hosts to secondary bacterial pneumonia, contributing to “excess mortality” that is observed during influenza epidemics and pandemics. During the 40 years that H3N2 viruses have circulated within humans, viruses belonging to this subtype have frequently been associated with “excess mortality,” and historical evidence demonstrates that Streptococcus pyogenes (group A streptococcus, GAS) is a key gram-positive pathogen that contributes to this secondary complication. Here we tested the hypothesis that vaccination against influenza virus can limit the development of influenza:GAS super-infections. Methods and Materials: To test this hypothesis, we established a model for influenza virus:GAS super-infection that involves administration of a sublethal dose of influenza virus expressing hemagglutinin (HA) from the A/Hong Kong/1/68 (HK68) H3N2 virus isolate in allantoic fluid (0.1 mouse 50% lethal dose, MLD50) one week prior to inoculation with a sublethal dose (0.1 MLD50) of a serotype M3 strain of GAS (MGAS 315). Results: Within our model, inoculation of groups with either allantoic fluid followed by saline (control, n = 6) or HK68 virus followed by PBS (virus alone, n = 18), was associated with 100% survival, while mice inoculated with allantoic fluid followed by MGAS 315 (bacteria alone, n = 22) demonstrated 95% survival. Alternatively, mice that were inoculated with HK68 virus followed by MGAS 315 (virus + bacteria, n = 19) demonstrated only 32% survival. After establishing a model of influenza virus:GAS super-infection, we tested our hypothesis that vaccination against influenza virus can limit secondary pneumonia. Specifically, we directly compared two influenza vaccines, inactivated influenza virus (IIV), and live, attenuated influenza virus (LAIV) for their ability to induce immunity in mice. Immune responses toward a single dose of vaccine yielded hemagglutination inhibition (HAI) titers of 145 ± 114 for IIV (n = 21) and 175 ± 48 for LAIV (n = 21). A boost with the respective vaccines yielded HAI titers of 725 ± 353 (IIV) and 448 ± 174 (LAIV). Antibody toward HK68 HA was not detected within the sera of control mice (n = 33) using the HAI assay. We challenged all of the mice with 0.1 MLD50 HK68 virus, followed one week later with 0.1 MLD50 MGAS 315 as detailed above. Within this super-infection model, we observed only a 16% survival for unvaccinated mice (n = 19), which was different from the survival of mice vaccinated with either IIV (67%, n = 9) or LAIV (78%, n = 9). Of note, 24 hours after inoculation with MGAS 315, bronchoalveolar lavage fluid (BALF) was collected from a subset of mice, and bacteria were observed microscopically within all lungs (n = 12, 4 per group). Surprisingly, GAS was cultured from BALF of vaccine control mice (4/4) and mice vaccinated with IIV (1/4), but not mice vaccinated with LAIV (0/4), suggesting a difference in viable bacteria within the lungs of vaccinated mice. Conclusion: We have demonstrated that vaccination against influenza virus, using either IIV or LAIV can elicit systemic antibody responses against influenza virus that limit the development of pneumonia in an influenza virus:GAS super-infection model. The observation that bacteria were present in BALF samples collected from all mice, but only consistently viable within the BALF from unvaccinated mice suggests, a difference in immunity within the lungs of vaccinated mice, likely due to contributions from influenza-specific antibodies that were present prior to virus challenge. We conclude that both IIV and LAIV can limit progression toward secondary complications in our model of influenza:GAS super-infection. Lungs collected 24 hours after MGAS 315 inoculation (day 8 post-influenza) were embedded in paraffin, sectioned, and stained with H&E (200X magnification) to examine the histopathology. At the same time, bacteria were cultured from the BALF collected from both the vaccine control (4/4) and the IIV (1/4) groups, but not from the LAIV group (0/4). Alternatively, bacteria were cultured from lung homogenates taken from mice in all groups (n = 4), with only a single mouse in the LAIV group demonstrating no bacteria in its lung (data not shown). Influenza virus was not detected in the lung homogenates of individual mice (n = 4) from any of the three groups. Sera collected 3 weeks after inoculation with IIV, LAIV, or vaccine control groups were analyzed for influenza-specific reactivity using a standard HAI assay. Antibody responses were enhanced after both primary and secondary inoculation against influenza virus, regardless of the vaccine used (IIV or LAIV). Of note, both vaccines induced similar increases in systemic (serum) antibody titers after both primary and secondary inoculations. Virus was created using the 8-plasmid reverse genetics system, incorporating plasmids expressing A/Puerto Rico/8/34 PB1, PB2, PA, M, NS, and NP genes and plasmids expressing H3N2 HA or NA genes (A/Hong Kong/1/68, HK68). A sub-lethal dose (0.1 MLD50) of HK68 HA-expressing influenza virus was administered to mice by the intranasal (i.n.) route 7 days prior to i.n. inoculation with a sub-lethal dose (0.1 MLD50) of a well-characterized, invasive isolate of Streptococcus pyogenes (Group A Strep, GAS) expressing serotype 3 M protein (MGAS 315). Weight loss (morbidity) and survival (mortality) are presented. Mice in all three vaccine groups were inoculated i.n. with influenza virus (0.1 MLD50) seven days prior to i.n. inoculation with MGAS 315 (0.1 MLD50). Mice were weighed daily and monitored for mortality. Twenty-four hours after inoculation with bacteria (Day 8 post-influenza challenge), mice were euthanized (n = 4) and BALF was collected by flushing the lungs with 1 mL PBS containing 5 mM EDTA. Analysis of specific antibody isotypes (IgG1, IgG2a, and IgA) within the BALF was completed using an ELISA (1 mg mL-1 HA for coating). IgA levels within the BALF of LAIV recipients were increased over the levels seen in the IIV and vaccine control groups, demonstrating induction of local (mucosal) immunity within these vaccinated animals. Surprisingly, the IgG1 and IgG2a responses in both IIV and LAIV recipients were not increased when compared to the IgG1 and IgG2a levels detected in the vaccine control group, despite the fact that both isotypes were elevated in serum prior to challenge (data not shown). Also prior to challenge, influenza-specific IgA in serum was not detected in any vaccine group (data not shown). ACKNOWLEDGMENTS USD Animal Resource Center Jonathan A. McCullers (St. Jude Children’s Research Hospital) for providing viruses SD-BRIN Undergraduate Fellows Program: NIH Grant Number 2 P20 RR016479 from the INBRE Program of the National Center for Research Resources (L.A.A.) USD U. Discover Undergraduate Scholarship Program (M.S. and F.P.D.) USD New Faculty Development Award (V.C.H.) USD-BBS New Faculty Start-up (V.C.H.)