Download

1 / 18
180 likes | 370 Views
The sequence structure of human nucleosome DNA. The team. Simon B. Kogan and Edward N. Trifonov; University of Haifa (Israel) Megumi Kato, Yoshiaki Onishi and Ryoiti Kiyama; National Institute of Advanced Industrial Science and Technology (Japan)

E N D
The team • Simon B. Kogan and Edward N. Trifonov;University of Haifa (Israel) • Megumi Kato, Yoshiaki Onishi and Ryoiti Kiyama;National Institute of Advanced Industrial Science and Technology (Japan) • Yuko Wada-Kiyama; Nippon Medical School (Japan) • Takashi Abe, Toshimichi Ikemura; National Institute of Genetics (Japan)
High-order chromatin • Chromatin is protein-DNA complex from which eukaryotic chromosomes are comprised (Euchromatin and heterochromatin). • Levels of chromatin organization: 300-nm loops; 100-nm fiber ?; 30-nm fiber; nucleosomes. • Models of 30-nm fiber: solenoid; zigzag; sallow thorn.
Electron Micrographs of Chromatin Isolated metaphase chromosome Modified from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, 12th edition, Figure 1-14 (BioMEDIA web site) From the web site of Dr. Carol Heckman (Bowling Green State University).
10-30 nm fibers and fibrils Modified from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, 12th edition, Figure 1-12 (BioMEDIA web site)
Levels of chromatin structure From The University of Edinburgh Faculty of Medicine web site
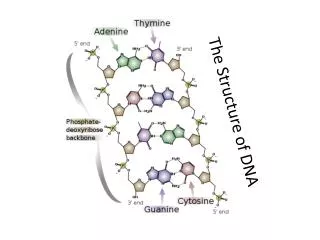
Nucleosome • Nucleosome is the basic block (lowest level) of chromatin organization. • Most of genomic DNA is confined in nucleosomes (one nucleosome per 200 base pairs on average). • Nucleosome core particle consist of histone octamer (two molecules each of H2A, H2B, H3 and H4 histone proteins) and stretch of super-coiled double-stranded DNA 125-166 base pairs long.
Nucleosome core particle Ribbon traces for the 146-bp DNA phosphodiester backbones (brown and turquoise) and eight histone protein main chains (blue: H3; green: H4; yellow: H2A; red: H2B (Luger et al., 1997).
Chromatin function • Package of DNA into chromosomes and nucleus. • Condense chromatin (heterochromatin) silences large chromosome DNA parts. • Epigenetic regulation of gene expression. • Low and high order chromatin structures are flexible and dynamic (i.e. chromatin remodeling). They regulate biological activity by hiding or exposure of DNA sites for protein binding.
Nucleosome function • Local higher-order chromatin structure depends on positions of individual nucleosomes. • The level of DNA sequence exposure to variety of binding factors depends on whether the sequence is constrained in nucleosome core or belongs to linker DNA. • Nucleosomes are positioned specifically in gene promoters and splice junctions. Thus, influence gene expression.
Nucleosome positioning • Nucleosome histones bind to DNA by means of electrostatic forces. However, the binding strength depends on specific DNA sequence. Thus, sequence can modulate nucleosome positioning preferences. • The important positioning factors are thought to be sequence’s anisotropic flexibility and curvature. Both of them depend mostly on base pairs interactions in dinucleotides. • The anisotropy can be achieved by periodical positioning of specific dinucleotides on the distance equal to DNA helix period in nucleosomes: ~10.4 bases.
Positioning pattern • Experimental identification of nucleosome positions is a cumbersome task and results are often imprecise and unreliable. Thus, the need for computational biology methods. • Because of nucleosome abundance, the positioning signal is necessary weak or very diverse to allow overlapping of other DNA codes (i.e. amino-acid triplet code). • However, several studies, observed weak AA(TT) dinucleotide periodicity (Trifonov and Sussman, 1980). Consequently, AA(TT) positioning pattern was built (Ioshikhes et al., 1996). More general RR(YY) pattern was built even earlier (Mengeritsky and Trifonov, 1983).
RR(YY) pattern in humans • Nobody (to the best of our knowledge) found the 10.4 periodicity in human genome. The presumed reason is the exceptional weakness of nucleosome preferential positioning in higher eukaryotes. • Utilizing large database of human dinucleosome DNA obtained from laboratory of Prof. Kiyama, we manage to obtain RR(YY) nucleosome positional periodical pattern (Kato et al., 2003) and get additional confirmation of nucleosome steric exclusion rules described earlier (Ulanovsky and Trifonov, 1986).
RR(YY) pattern A symmetrized sum of RR and YY distributions. The extrema corresponding to the 10.4*n ladder are indicated by arrows. Nucleosome sequences were smoothed by running the average of three positions (Kato et al., 2003).
GG(CC) pattern in humans • Detailed investigation of database sequences revealed predominant usage of GG(CC) dinucleotides in humans. • GG(CC) predominance is in contradiction with the current opinion that only AA(TT) and TA are important for nucleosome positioning. • This finding points to possible species specificity of nucleosome positioning signal. • The result was confirmed in independent study concerning the connection between nucleosome and splicing junction positions (Kogan and Trifonov, 2005).
Weights of eight topmost periodical components of splice junction dinucleotide profiles in four species (Kogan and Trifonov, 2005) Second line indicates amount of EI(IE) splice junctions of the respective species in the data set. For the purpose of comparison, the weights are calculated as fitting sine amplitudes divided by the sum of all 32 amplitudes, for each species separately.
Selected references • Bolshoy, A.: CC dinucleotides contribute to the bending of DNA in chromatin. Nat Struct Biol 2 (1995) 446-8. • Herzel, H., Weiss, O. and Trifonov, E.N.: 10-11 bp periodicities in complete genomes reflect protein structure and DNA folding. Bioinformatics 15 (1999) 187-93. • Ioshikhes, I., Bolshoy, A., Derenshteyn, K., Borodovsky, M. and Trifonov, E.N.: Nucleosome DNA sequence pattern revealed by multiple alignment of experimentally mapped sequences. J Mol Biol 262 (1996) 129-39. • Kato, M., Onishi, Y., Wada-Kiyama, Y., Abe, T., Ikemura, T., Kogan, S., Bolshoy, A., Trifonov, E.N. and Kiyama, R.: Dinucleosome DNA of human K562 cells: experimental and computational characterizations. J Mol Biol 332 (2003) 111-25. • Kogan, S. and Trifonov, E.N.: Gene splice sites correlate with nucleosome positions. GENE, in press (2005). • Mengeritsky, G. and Trifonov, E.N.: Nucleotide sequence-directed mapping of the nucleosomes. Nucleic Acids Res 11 (1983) 3833-51. • Thastrom, A., Lowary, P.T., Widlund, H.R., Cao, H., Kubista, M. and Widom, J.: Sequence motifs and free energies of selected natural and non-natural nucleosome positioning DNA sequences. J Mol Biol 288 (1999) 213-29. • Trifonov, E.N. and Sussman, J.L.: The pitch of chromatin DNA is reflected in its nucleotide sequence. Proc Natl Acad Sci U S A 77 (1980) 3816-20.