Download

1 / 49
550 likes | 1.55k Views
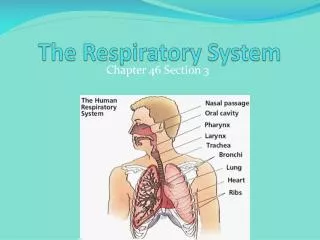
The avian respiratory system . And Respiration. House sparrow. The avian respiratory system delivers oxygen from the air to the tissues and also removes carbon dioxide . In addition, the respiratory system plays an important role in thermoregulation (maintaining normal body temperature).

E N D
The avian respiratory system And Respiration
The avian respiratory systemdelivers oxygen from the air to the tissues and also removes carbon dioxide. In addition, the respiratory system plays an important role in thermoregulation (maintaining normal body temperature). • The avian respiratory system is different from that of other vertebrates, with birds having relatively small lungs plus nine air sacs that play an important role in respiration (but are not directly involved in the exchange of gases).
Most birds have 9 air sacs:one interclavicular sac, • twocervical sacs ,two anterior thoracic sacs , • two posterior thoracic sacs ,two abdominal sacs. Functionally, these 9 air sacs can be divided into anterior sacs, or Cranial(interclavicular , cervicals , & anterior thoracics) & posterior sacs, or Caudal(posterior thoracics & abdominals). Air sacs have very thin walls with few blood vessels. So, they do not play a direct role in gas exchange. Rather, they act as a 'bellows' to ventilate the lungs .
Avian respiratory system hd = humeral diverticulum of the clavicular air sac; adapted from Sereno et al. 2008)
Birds can breathe through the mouth or the nostrils (nares). Air entering these openings (during inspiration) passes through the pharynx & then into the trachea (or windpipe). The trachea is generally as long as the neck.However, some birds, such as cranes(كركى - غرنوق), have an exceptionally long (up to 1.5 m) trachea that is coiled within the hollowed keel of the breast bone (shown below). This arrangement may give additional resonance to their loud calls.
The typical bird trachea is 2.7 times longer and 1.29 times wider than that of similarly-sized mammals. The net effect is that tracheal resistance to air flow is similar to that in mammals, but the tracheal dead space volume is about 4.5 times larger. Birds compensate for the larger tracheal dead space by having a relatively larger tidal volume and a lower respiratory frequency,approximately one-third that of mammals. These two factors lessen the impact of the larger tracheal dead space volume on ventilation. Thus, minute tracheal ventilation is only about 1.5 to 1.9 times that of mammals.
The trachea bifurcates (or splits) into two primary bronchi at the syrinx. The syrinx is unique to birds & is their 'voicebox' (in mammals, sounds are produced in the larynx). The primary bronchi(extrapulmonary)enter the lungs & are then called mesobronchi(intrapulmonary). Branching off from the mesobronchi are smaller tubes called dorsobronchi(mediodorsal sec.b.). The ventrobronchi( medioventral sec.b), in turn, lead into the still smaller parabronchi.
Parabronchi can be several millimeters long and 0.5 - 2.0 mm in diameter (depending on the size of the bird) and their walls contain hundreds of tiny, branching, & anastomosing 'air capillaries' surrounded by a profuse network of blood capillaries. Birds exhibit some variation in lung structure and, specifically, in the arrangement of parabronchi.Most birds have two sets of parabronchi, the paleopulmonic (‘ancient lung’) and neopulmonic (‘new lung’) parabronchi. However, the neopulmonic region is absent in some birds (e.g., penguins) and poorly developed in others (e.g.,ducks[Anatidae]). ]). In songbirds (Passeriformes), pigeons (Columbiformes), and gallinaceous birds (Galliformes), the neopulmonic region of the lung is well-developed.
Avian respiratory system structure Trachea 2 Extrapulmonary primary bronchi 2 Intrapulmonary primary bronchi Abdominal Air sac Mesoventral Scab. Mediodorsal Sec.b. Lateroventral Sec.b. (Caudal air sac) Parabronchi Parabronchi Join Paleopulmonic parabronchi Cervical air sac Clavicular air sac Cranial air sac In some species there is a Laterodorsal Sec.b. from which arises a neopulmonary parabronchi. The laterodorsal bronchi have connection to clavicular and Cranial air sacs.
Differences among different birds in the development of the neopulmonic region of the lung. (a) Penguin lungs are entirely paleopulmonic.
(b) Some birds, such as ducks, have a relatively small neopulmonic region.(c) Songbirds have a well-developed neopulmonic region.
Embryological Development • The first appearance of the lungs of the chick embryo comes in the early part of the third day of development. At their beginning , the primitive lungs are paired and consist of two shallow pouches that open widely into the floor of the pharynx. • The bronchial system developed from the lung pouches. • At the end of the fourth day of incubation , the trachea becomes differentiated from the caudal portion of the laryngotracheal groove, and at the distal end of the lung tube there is an enlargement that later becomes the abdominal air sac.
The secondary bronchi begin to develop in the sixth day. • On the ninth day the lung has increased in size dorsoventrally and occupies a more lateral position in the thoracic cavity . The five air sacs are formed and project beyond the surface of the lung. • On the tenth day, outgrowths from the abdominal and caudal thoracic air sacs appear called “recurrent bronchi’ • ,which eventually ramify and anastomose with parabronchi in various parts of the lung. At 10 ½ days of incubation, similar outgrowths appear from all air sacs except the cervical sac. The parabronchi are fully formed by the eighteenth day of incubation , and the air capillaries are fully developed between the nineteenth and twenty-first days of development.
Respiration by Avian Embryos During avian development there are three sequential stages of respiration : prenatal (embryonic), paranatal (hatching), and postnatal (post hatching). During the prenatal stage respiratory gas exchange occurs via diffusion between the external environment and the initial gas exchanger (i.e., the area vasculosa, or the region of blood island formation and forerunner of the chorioallantoic membrane) in early embryonic life and later the vascular bed of the chorioallantois.
The paranatal stage starts when the beak penetrates into the air pocket (air cell) between the inner and outer shell membranes (both internal to shell; i.e., internal pipping) this occurs during the last 2-3 days of incubation. During this stage, the lungs begin to replace the chorioallantois as the gas exchanger, yet diffusion remains the major mechanism moving gas across the shell. The postnatal stage begins when the beak penetrates the shell (i.e., external pipping) .
The shell is pierced by a great number of fine pores which are filled by an organic(protein) substance related to collagen. In an average hen’s egg the pores have a diameter of 0.04 to 0.05 mm ., and the total number is estimated at about 7000.
Mechanics of breathing.Forces that move gases through the lung. • The forces required to move gas through the lungs are derived from the action of the respiratory muscles. • Codd et al. (2005) examined the activity of three musclesassociated with the uncinate processes, (1) external intercostal, (2)appendicocostalis and (3) external oblique (labeled in drawing to the left) examined using electrodes during sitting, standing and moderate speed treadmill running in a Giant Canada Goose.
The external intercostal muscles demonstrated no respiratory activity, being active only during running, suggesting they play some role in trunk stabilization. Theappendicocostalisand external oblique muscles are respiratory muscles, being active during inspiration and expiration, respectively. The activity of the appendicocostalis muscle increased when sternal movements were restricted, suggesting that activity of these muscles may be particularly important during prolonged sitting such as during egg incubation
Codd et al. (2005) suggested that the uncinate processes in birds facilitate movements of the ribs and sternum during breathing and therefore are integral to the breathing mechanics of birds.
*Changes in the position of the thoracic skeleton during breathing in a bird. The solid lines represent thoracic position at the end of expiration while the dotted lines show the thoracic position at the end of inspiration
Pattern and direction of gas flow through the avian lung. • The avian pulmonary system uses "flow-through ventilation," relying on a set of nine flexible air sacs that act like bellowsto move air through the almost completely rigid lungs. Air sacs do not take part in the actual oxygen exchange, but do greatly enhance its efficiency and allow for the high metabolic rates found in birds. The movement of gas within the lungs of various species of birds has been measured directly .
The measurements show unequivocally that during both inspiration and expiration gas moves unidirectionallyfrom the intrapulmonary primary bronchus to the mediodorsal secondary bronchi and into the parabronchi of the paleopulmonic. However, gas moves bidirectionally through the neopulmonic and through the direct connections to the caudal air sacs(caudal thoracic and abdominal sacs) during both inspiration and expiration.
Part of the gas that reaches the caudal air sacs during inspiration passes through the neopulmonic parabronchi and is thereby exposed to gas-exchange surfaces . Such gas movement is responsible for the lower oxygen concentration and higher carbon dioxide concentration in the cranial air sacs compared with the caudal air sacs.
During inspiration: • the sternum moves forward and downward while the vertebral ribs move cranially to expand the sternal ribs and the thoracoabdominal cavity. This expands the posterior and anterior air sacs and lowers the pressure, causing air to move into those air sacs. Air from the trachea and bronchi moves into the posterior air sacs and, simultaneously, • Air from the lungs moves into the anterior air sacs. Gas that reaches the cranial air sacs must pass through the paleopulmonic parabronchi and undergo exchange with the blood .
Gas flow through Avian lung INSPIRATION INTRAPULMONARY AIR TRACHEA EXTRAPULMONARY P.B PART TO PART TO PART TO Paleopulmonic Directly to abdominal air sacs(caudal) (Cranial air sacs) Neopulmonic • Lower oxygen • Higher carbon dioxide • As compared to caudal air sacs ( Caudal air sacs)
During expiration On expiration , gas passes simultaneously from all air sacs to the trachea ,but over different pathways. Gas from the caudal air sacs passes through the neopulmonic in the opposite direction from that in inspiration ;part of it then moves into the mediodorsal bronchi ,through the paleopulmonic parabronchi ,through the medioventral bronchi to the intrapulmonary primary bronchus ,and out the trachea
EXPIRATION Cranial air sacs Caudal air sacs Medioventral Sec.b Neopulmonic P.b. Intrapulmonic P.b. Paleopulmonic Intrapulmonic P.b. Extrapulmonic P.b. Intrapulmonic P.b. Extrapulmonic P.b. Extrapulmonic P.b. TRACHEA TRACHEA No Exchange of GAS TRACHEA
Exchange of gases: • - In the avian lung, oxygen diffuses (by simple diffusion) from the 'air capillaries' into the blood & carbon dioxide from the blood into the 'air capillaries‘. • - Air (passing through the parabronchi) & blood (moving through capillaries) travel at right angles to each other. This is referred to as cross-current flow. Cross-current exchange is very efficient, with oxygen & carbon dioxide pressure gradients maintained along the entire length of the parabronchus-capillary 'connection.'
Top: Air flow (large arrows) and blood flow (small arrows) illustrating the cross-current gas-exchange mechanism operating in the avian lung (between the blood capillaries and air capillaries). Note the serial arrangement of blood capillaries running from the periphery to the lumen of the parabronchus and the air capillaries radially extending from the parabronchial lumen. The exchange of gases (simple diffusion of O2 and CO2) occurs only between blood capillaries and air capillaries. As air moves through a parabronchus and each successive air capillary, the partial pressure of oxygen (PO2) declines (as indicated by the decreased density of the stippling) because oxygen is diffusing into the blood capillaries associated with each air capillary.
As a result of this diffusion, the partial pressure of oxygen in the blood leaving the lungs (pulmonary vein) is higher than that in blood entering the lungs (pulmonary artery) (as indicated by the increased density of the stippling). Bottom: Relative partial pressures of O2 and CO2 (1) for air entering a parabronchus (initial-parabronchial, PI) and air leaving a parabronchus (end-parabronchial, PE), and (2) for blood before entering blood capillaries in the lungs (pulmonary artery, PA) and for blood after leaving the blood capillaries in the lungs (pulmonary vein, PV).
The partial pressure of oxygen (PO2) of venous blood (PV) is derived from a mixture of all serial air capillary-blood capillary units. Because of this cross-current exchange the partial pressure of oxygen in avian pulmonary veins (PV) is greater than that of the air leaving the parabronchus (PE); air that will be exhaled.
10 100 0 0 30 20 60 50 80 70 100 90 50 50 Simple exchange Counter-current exchange Examples :Exchange surface /unit body weight. Goose :17.9 Cm2 / g Pigeon:40.3 Cm2 / g Exchange surface / unit volume of lung. Turkey: 188 mm2 /mm2Pigeon:302 mm2 /mm2
Gas –exchange efficiency Blood Exit. Medium enter P O2 Blood enter Medium exit. Alveolar Uniform(M) Crosscurrent(B) Countercurrent (F)
Control of Ventilation: • Ventilation and respiratory rate are regulated to meet the demands imposed by changes in metabolic activity (e.g., rest and flight) as well as other sensory inputs (e.g., heat and cold). There is likely a central respiratory control center in the avian brain, but this has not been unequivocally demonstrated.
As in mammals, the central control area appears to be located in the pons and medulla oblongata with facilitation and inhibition coming from higher regions of the brain. It also appears that the chemical drive on respiratory frequency and inspiratory and expiratory duration depend on feedback from receptors in the lung as well as on extrapulmonary chemoreceptors, mechanoreceptors,and thermoreceptors (Ludders 2001).
Central chemoreceptors affect ventilation in response to changes in arterial PCO2 and hydrogen ion concentration.Peripheral extrapulmonary chemoreceptors, specifically the carotid bodies (located in the carotid arteries), are influenced by PO2 and increase their discharge rate as PO2 decreases, thus increasing ventilation; they decrease their rate of discharge as PO2 increases or PCO2 decreases. These responses are the same as those observed in mammals.
Unlike mammals, birds have a unique group of peripheral receptors located in the lung called intrapulmonary chemoreceptors (IPC) that are acutely sensitive to carbon dioxide and insensitive to hypoxia. The IPC affect rate and volume of breathing on a breath-to-breath basis by acting as the afferent limb of an inspiratory-inhibitory reflex that is sensitive to the timing, rate, and extent of CO2 washout from the lung during inspiration (Ludders 2001).