Download

1 / 65
710 likes | 1.77k Views
Chapter 18 Glycolysis. Outline. What are the essential features of glycolysis ? Why are coupled reactions important in glycolysis ? What are the chemical principles and features of the first phase of glycolysis ?

E N D
Outline • What are the essential features of glycolysis ? • Why are coupled reactions important in glycolysis ? • What are the chemical principles and features of the first phase of glycolysis ? • What are the chemical principles and features of the second phase of glycolysis ? • What are the metabolic fates of NADH and pyruvate produced in glycolysis ? • How is glycolysis regulated ? • Is glucose the only substrates used in glycolysis ?
Terms Used in Metabolism Pathway Steps Metabolites Glycolysis 10 glucose to pyruvate Anaerobic glycolysis 11 glucose to lactate Alcoholic Fermentation 12 glucose to ethanol/CO2 Respiration (Aerobic) - glucose to CO2 and HOH, (includes glycolysis, PDH, Krebs, ETS/OP) Substrate level phosphorylation: Synthesis of of ATP using energy from a metabolic reaction. Oxidative phosphorylation: Synthesis of ATP using energy from electron transport.
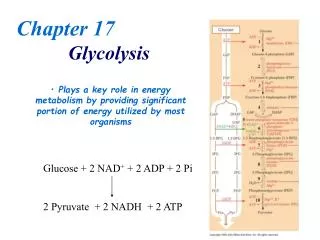
18.1 What are the Essential Features of Glycolysis ? Glycolysis is also called the Embden-Meyerhof Pathway • Essentially all cells carry out glycolysis. • Glycolysis consists of ten reactions which are essentially the same in all cells. These are divided into two phases. • The first phase converts glucose to two G-3-P and is the energy requiring phase. • The second phase generates two pyruvates and is the energy producing phase. • Products of glycolysis are pyruvate, ATP and NADH. • There are three major fates for pyruvate.
Figure 18.1 The Glycolytic Pathway Energy requiring Energy producing
The Fates of Pyruvate From Glycolysis Figure 18.2 Pyruvate produced in glycolysis can be utilized by cells in several ways. In animals, pyruvate is normally converted to acetyl-coenzyme A, which is then oxidized in the TCA cycle to produce CO2. When oxygen is limited, pyruvate can be converted to lactate. Alcoholic fermentation in yeast converts pyruvate to ethanol and CO2.
18.2 Why Are Coupled Reactions Important in Glycolysis? • Coupled reactions involving ATP hydrolysis are used to drive the glycolytic pathway. • Coupled reactions convert some, but not all of the metabolic energy of glucose into ATP. • Under cellular conditions, approximately 5% of the energy of glucose is released in glycolysis.
Steady-State Concentrations of Glycolytic Intermediates These steady-state concentrations are used to obtain the cellular values of ΔG found in Table 18.1 and Figure 18.22.
Rxn 1: Hexokinase The first reaction - phosphorylation of glucose • Catalyzed by hexokinase or glucokinase. • This is a priming reaction - ATP is consumed here in order to get more later. • ATP makes the phosphorylation of glucose thermodynamically spontaneous. • Be sure you can interconvert Keq and standard state free energy change. • Be sure you can use Eq. 3.13 to generate far right column of Table 18.1.
Calculation of the overall ΔG for Rx 1 Glucose + ATP <==> Glucose-6-P + ADP G = Go’ + RT ln (G-6-P)(ADP)/ (Glucose)(ATP) (0.083x10-3)(0.83x10-3) G = -16700 + RT ln ------------------------------ (5x10-3)(1.85x10-3) G= -16700 + 2577 ln (1.256x10-3) = -16700 + 2577 (-6.68) = -16700 – 17213 = -33910 or -33.91 kJ/mol
Rxn 1: Hexokinase • G for the 1st step is large and negative. • Phosphorylation of glucose by hexokinase is done to keep glucose in the cell. • Glucose-6-P has no transporter and is trapped. • Km for glucose is 0.1 mM; cell has 4 mM glucose. • So hexokinase is normally active! • Glucokinase (Kmglucose = 10 mM) only turns on when cell is rich in glucose. • Hexokinase is regulated - allosterically inhibited by (product) glucose-6-P - but is not the most important site of regulation of glycolysis - Why?
Glucose is kept in most cells by phosphorylation to glucose-6-phosphate Figure 18.4 Glucose-6-P cannot cross the plasma membrane.
Glucose-6-P, intermediate forming an important branch point in metabolism Figure 18.5 Glucose-6-phosphate is the branch point for several metabolic pathways.
Rxn 1: Hexokinase Figure 18.6 The (a) open and (b) closed states of yeast hexokinase. Binding of glucose (green) induces a conformation change that closes the active site, as predicted by Daniel Koshland. The induced fit model for enzymes is discussed on page 409 of the text.
Rxn 2: Phosphoglucoisomerase Glucose-6-P to Fructose-6-P • Why does this reaction occur ? • next step (phosphorylation at C-1) would be tough for hemiacetal -OH, but easy for primary –OH. • isomerization activates C-3 for cleavage in aldolase reaction. • An ene-diol is an intermediate in this reaction . • Be able to write a mechanism for this interesting enzyme reaction (see Figure 18.8).
A mechanism for phosphoglucoisomerase Figure 18.8 The phosphoglucoisomerase mechanism involves opening of the pyranose ring (step 1), proton abstraction leading to enediol formation (step 2), and proton addition to the double bond, followed by ring closure (step 3)
Rxn 3: Phosphofructokinase (PFK-1) PFK-1 catalyzes the committed step of glycolysis • The second priming reaction of glycolysis. • It is the committed step, has a large, negative ΔG and PFK is highly regulated (key control point). • ATP inhibits, AMP reverses inhibition. • Citrate is also an allosteric inhibitor. • Fructose-2,6-bisphosphate is allosteric activator. • PFK-1 increases activity when energy status is low. • PFK-1 decreases activity when energy status is high.
Phosphofructokinase – a Second Phosphorylation Driven by ATP Phosphofructokinase is the second energy requiring reaction of glycolysis. ATP is consumed in this priming reaction, so that more ATP can be produced further along the pathway. PFK-1 produces fructose-1,6-bisphosphate, a glycolytic intermediate. PFK-2 produces fructose-2,6-bisphosphate, a regulator of PFK-1.
PFK-1 behaves cooperatively at high [ATP] Figure 18.9 At high ATP, phosphofructokinase (PFK) behaves cooperatively and the activity plot is sigmoid.
PFK-1 is activated by F-2,6-BP Figure 18.10 Fructose-2,6-bisphosphate (F-2,6-BP) activates PFK, increasing the affinity of the enzyme for fructose-6-phosphate and restoring the hyperbolic dependence of enzyme activity on substrate concentration.
F-2,3-BP effect on inhibition of PFK by ATP Figure 18.11 F-2,6-BP stimulates PFK by decreasing the inhibitory effects of ATP. i.e. F-2,6-BP overrides the effect of ATP.
Effect of AMP on PFK-1 and F-1,6-BisPase PFK-1 and fructose-1,6-bisphosphatase are reciprocally regulated. When one is turned on the other is turned off and visa versa. However, the activity reached by PFK-1 is greater that of the bisphosphatase due to the greater amount of PFK-1 present in the cell. The bisphosphatase will be discussed more later.
Rxn 4: Aldolase • A C6 intermediate is cleaved to 2 C3s (DHAP and Gly-3-P). • Animal aldolases are Class I aldolases. • In Class I aldolases, a covalent Schiff base intermediate is formed between the substrate and an active site lysine . • The evidence for a Schiff base intermediate for Class I aldolases is described in Problem 18 on page 562.
The fructose biophosphate aldolase Rxn The aldolase reaction is unfavorable as written at standard state. The cellular ΔG, however, is close to zero.
Figure 18.12 Mechanism for the Class I aldolase reaction, showing the Schiff base as electron sink
A Class II aldolase mechanism Figure 18.12(b) In Class II aldolases, an active-site Zn2+ stabilizes the enolate intermediate, leading to polarization of the substrate carbonyl group.
Rxn 5: Triose Phosphate Isomerase Triose phosphate isomerase completes the first phase of glycolysis. Each glucose has been converted to two molecules of glyceraldehyde-3-phosphate.
Rxn 5: Triose Phosphate Isomerase • DHAP is converted to G-3-P. • This reaction makes it possible for both products of the aldolase reaction to continue in glycolysis. • The reaction involves an ene-diol mechanism. • Glu165 in the active site acts as a general base. • Triose phosphate isomerase is a near-perfect enzyme - see Table 13.5.
Mechanism for triose phosphate isomerase Figure 18.13 A reaction mechanism for triose phosphate isomerase. In the yeast enzyme, the catalytic residue is Glu165.
18.4 What Are the Chemical Principles and Features of the Second Phase of Glycolysis? • Metabolic energy of glucose produces 4 ATP. • Net ATP yield for glycolysis is two ATP. • The second phase of glycolysis involves two very high energy phosphate intermediates: • 1,3-bisphosphoglycerate. • Phosphoenolpyruvate.
Rxn 6: Glyceraldehyde-3-P Dehydrogenase G-3-P is oxidized to 1,3-BPG • Energy yield from converting an aldehyde to a carboxylic acid is used to make 1,3-BPG and NADH. • The mechanism involves covalent catalysis and a nicotinamide coenzyme, and it is good example of nicotinamide chemistry. • This enzyme reaction is the site of action of arsenate – an anion analogous to phosphate.
The glyceraldehyde-3-phosphate dehydrogenase reaction The only reaction of glycolysis that requires Pi and NAD+.
Rxn 6: Glyceraldehyde-3-P Dehydrogenase Figure 18.14 A mechanism for the glyceraldehyde-3-phosphate dehydrogenase reaction. Reaction of an enzyme sulfhydryl with G3P forms a thiohemiacetal, which loses a hydride to NAD+ to become a thioester. Phosphorolysis releases 1,3-bisphosphoglycerate.
G3P-DH is the site of action of arsenate If arsenate is used as substrate for the G3P-DH reaction, it forms 1-arseno-3-phosphoglycerate. This product spontaneously breaks down to 3-phosphoglycerate, which bypasses the phosphoglycerate kinase reaction. The result is that glycolysis in the presence of arsenate produces no net ATP.
Rxn 7: Phosphoglycerate Kinase ATP synthesis from a high-energy phosphate • Phosphoglycerate kinase transfers a phosphoryl group from 1,3-bisphosphoglycerate to ADP to form an ATP. • This is referred to as “substrate-level phosphorylation”. • This reaction “pays off” the ATP debt created by the priming reactions in the first phase.
Rxn 7: Phosphoglycerate Kinase Phosphoglycerate kinase transfers a phosphoryl group from 1,3-bisphosphoglycerate to ADP to form an ATP. This is referred to as a “substrate-level phosphorylation”.
Rxn 7: Phosphoglycerate Kinase The open (a) and closed (b) forms of phosphoglycerate kinase. ATP (cyan), 3-phosphoglycerate (purple), and Mg2+ (gold).
Rxn 8: Phosphoglycerate Mutase Phosphoglycerate mutase catalyzes a phosphoryl group transfer from C-3 to C-2 • Rationale for this reaction in glycolysis: It repositions the phosphate to make PEP in the following reaction (enolase). • Note the phospho-histidine intermediates. • Zelda Rose (wife of Nobel laureate Irwin Rose) showed that a trace of 2,3-BPG is required to phosphorylate His. • Nomenclature note: a “mutase” catalyzes migration of a functional group within a substrate.
Rxn 8: Phosphoglycerate Mutase Figure 18.17 A mechanism for the phosphoglycerate mutase reaction in rabbit muscle and in yeast. Zelda Rose showed that the enzyme requires a small amount of 2,3-BPG to phosphorylate the His residue before the mechanism can proceed.
2,3-BPG synthesis in erythrocytes detours around the phosphoglycerate kinase rxn Most cells contain only a trace of 2,3-BPG, but erythrocytes typically contain 4-5 mM 2,3-BPG. 2,3-bisphosphoglycerate is an important regulator of hemoglobin (see pages 475-476 of the text). 2,3-BPG (for hemoglobin) is made by circumventing the PGK reaction (Figure 18.15). 2,3-BPG is formed from 1,3-BPG by bisphosphoglycerate mutase. 3-phosphoglycerate is then formed by 2,3-bisphosphoglycerate phosphatase.
Erythrocyte synthesis of 2,3-BPG Figure 18.15 Formation and decomposition of 2,3-bisphosphglycerate in erythrocytes (not glycolysis). This synthesis does not result from a direct internal transfer but requires 3-phosphoglycerate as substrate. There is no ATP formed in this conversion.
Rxn 9: Enolase Conversion of 2-Phosphoglycerate to PEP • The enolase makes a high-energy phosphate in preparation for ATP synthesis in step 10. • The overall Go’ for this reaction is +1.8 kJ/mol. • How can such a reaction create a PEP ? • Enolase just rearranges 2-PG to a form from which more energy can be released in hydrolysis. • Fluoride ion inhibits enolase.
Rxn 9: Enolase The enolase reaction creates a high-energy phosphate in preparation for ATP synthesis in step 10 of glycolysis.
Rxn 10: Pyruvate Kinase Conversion of PEP to pyruvate makes ATP • These two ATP (from one glucose) can be viewed as the "payoff" of glycolysis. • Large, negative G – indicating that this reaction is subject to allosteric regulation. • PK is activated by AMP, F-1,6-bisP. • PK is inhibited by ATP and Acetyl-CoA. • Understand the keto-enol equilibrium of pyruvate; it is the key to understanding the pyruvate kinase reaction.
Rxn 10: Pyruvate Kinase The pyruvate kinase reaction converts PEP to pyruvate, driving synthesis of ATP. The two ATP produced here from one glucose are the “payoff” of glycolysis.
Rxn 10: Pyruvate Kinase Figure 18.19 The conversion of phosphoenolpyruvate (PEP) to pyruvate may be viewed as involving two steps: phosphoryl transfer, followed by an enol-keto tautomerization. The tautomerization is spontaneous and accounts for much of the free energy change for PEP hydrolysis.
18.5 What Are the Metabolic Fates of NADH and Pyruvate Produced in Glycolysis? NADH can be recycled via aerobic or anaerobic pathways • NADH is energy - two possible fates: • In aerobic conditions (O2 is available), NADH is re-oxidized in the electron transport pathway, making ATP in oxidative phosphorylation. • In anaerobic conditions, NADH is re-oxidized by lactate dehydrogenase (LDH) or alcohol dehydrogense either of which provide NAD+ for more glycolysis.