Download
1 / 109
1.24k likes | 2.75k Views
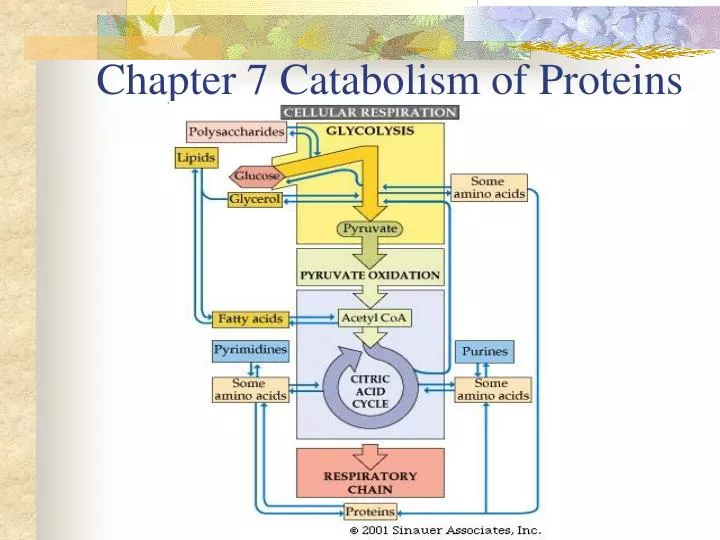
Chapter 7 Catabolism of Proteins. Nutritional Function of Proteins. Functions : Structural Catalytic, Transport action Signaling and hormonal functions Source of energy (16.7kJ/g). Nutritional Requirement of Proteins.

E N D
Nutritional Function of Proteins Functions: Structural Catalytic, Transport action Signaling and hormonal functions Source of energy (16.7kJ/g)
Nutritional Requirement of Proteins Nitrogen Balance Proteins contain about 16% nitrogen Intake N = losses N Intake N > Losses N Intake N < Losses N
Nutritional Quality of Proteins Essential Amino Acids cannot be synthesized by the body and must be obtained from diet Eight nutritional essential amino acids Tryptophan Phenylalanine Lysine Threonine Valine Leucine Isoleucine methionine
Nutritional Quality of Proteins Non-essential amino acids synthesized in the body synthesized by the transamination of a-keto acids Tyrosine and cysteine synthesized in the body by using essential amino acids from phenylalanine and methionine respectively semi-essential
Digestion of Dietary Proteins Dietary proteins are digested in the stomach and intestine
Digestion of Protein in the Stomach The digestion of protein. Protein is broken down into amino acids by the enzymes pepsin (secreted by the stomach) and trypsin and peptidase (in the small intestine).
Table 1. Phases of Digestion and Absorption of Protein and its Degradative Products
carbonic anhydrase CO2 + H2O H2CO3 H+ H+ HCO3- HCO3- K+ ATP ADP + Pi Cl- Cl- Cl- H+,K+-ATPase Gastric Parietal Cell Lumen of the Stomach Plasma CO2 Production of gastric acid and its secretion
autoactivation (intramolecular cleavage) Pepsin aa Dietary Protein Phase 1- Gastric digestion Figure 2. Gastric digestion of dietary protein. Gastric Chief Cells Pepsinogen denaturation by stomach acid hydrolysis by pepsin autocatalysis large peptide fragments free amino acids aa aa Pyloric sphincter aa Duodenum • Acid from parietal cells denatures protein to be more susceptible to pepsin cleavage. • Pepsinogen activated to pepsin by autoactivation and autocatalysis by pepsin. • Large peptide fragments/some amino acids pass through the pyloric sphincter to the duodenum
free amino acids from gastric digestion Entero-peptidase Trypsin Phase 2- Digestion by pancreatic proteases Duodenal Endocrine Cell Duodenal Endocrine Cell CCK-PZ CCK-PZ Trypsinogen Blood- stream (hydrolysis) Pancreatic Acinar Cell Mucosal Epithelial Cells Figure 3. Secretion, activation and action of pancreatic proteases and brush border endopeptidases and aminopeptidases
Phase 2- Digestion by pancreatic proteases Duodenal Endocrine Cell CCK-PZ Duodenal Endocrine Cell CCK-PZ free amino acids from gastric digestion Trypsinogen Secretin Entero-peptidase (hydrolysis) Blood- stream autocatalysis Trypsin Pancreatic Acinar Cell HCO3- neutralizes acid Mucosal Epithelial Cells Figure 3. Secretion, activation and action of pancreatic proteases and brush border endopeptidases and aminopeptidases
catalysis Chymotrypsin Elastase Carboxypeptidases Phase 2- Digestion by pancreatic proteases Duodenal Endocrine Cell CCK-PZ Duodenal Endocrine Cell CCK-PZ Secretin free amino acids from gastric digestion Trypsinogen Entero-peptidase (hydrolysis) Blood- stream autocatalysis Trypsin Pancreatic Acinar Cell Chymotrypsinogen Proelastase Procarboxypeptidases HCO3- neutralizes acid Mucosal Epithelial Cells Figure 3. Secretion, activation and action of pancreatic proteases and brush border endopeptidases and aminopeptidases
Figure 3. Secretion, activation and action of pancreatic proteases and brush border endopeptidases and aminopeptidases Phase 2- Digestion by pancreatic proteases Phase 3- Digestion at the brush border Duodenal Endocrine Cell CCK-PZ Duodenal Endocrine Cell CCK-PZ Secretin free amino acids from gastric digestion Trypsinogen Entero-peptidase (hydrolysis) Blood- stream autocatalysis Trypsin Pancreatic Acinar Cell Chymotrypsinogen Proelastase Procarboxypeptidases catalysis HCO3- neutralizes acid Mucosal Epithelial Cells Chymotrypsin Elastase Carboxypeptidases amino acids dipeptides tripeptides brush border endo-/aminopeptidases hydrolyze products; amino acids, di-/tripeptides absorbed by epithelial cells
Di-, tri- peptides Dipeptides, tripeptides Dipeptidases, tripeptidases 2K+ 3Na+ ATP Amino acids ADP + Pi 3Na+ 2K+ = Na+-dependent co-transport = Na+,K+-ATPase LUMEN OF INTESTINE Na+ Amino acids Intestinal Epithelium Phase 4 - Absorption Brush border Na+ contraluminal membrane Figure 4. Absorption of amino acids and di- and tripeptides from the intestinal lumen
BRUSH BORDER TRANSPORT SYSTEMS a) neutral amino acids (uncharged aliphatic and aromatic) b) basic amino acids and cystine (Cys-Cys) c) acidic amino acids (Asp, Glu) d) imino acids (Pro) e) dipeptides and tripeptides
Di-, tri- peptides Dipeptidases, tripeptidases 2K+ 3Na+ ATP ADP + Pi 3Na+ 2K+ = Na+-dependent co-transport = Na+,K+-ATPase = facilitated diffusion Na+ Amino acids LUMEN OF INTESTINE Phase 4 - Absorption Intestinal Epithelium Dipeptides, tripeptides Brush border Phase 5 Amino acids contraluminal membrane Phase 5 capillaries Figure 4. Absorption of amino acids and di- and tripeptides from the intestinal lumen
Putrefaction Decomposition of amino acids and proteins by bacteria Most ingested proteins are absorbed from the small intestine 95% of total dietary proteins Undigested proteins pass into the large intestine Bacterial activity occurs
Putrefaction Bacteria putrefaction produces some nutritional benefits, Vitamin K, Vitamin B12, Folic acid Toxic for human Amines, phenol, indole, H2S
Production of Amines • Production of phenol • Production indole • Production of H2S • Production of Ammonia Page 209
dC Rate of Turnover = = KS - KDC dt The half-life of proteins is determined by rates of synthesis and degradation A given protein is synthesized at a constant rate KS A constant fraction of active molecules are destroyed per unit time KS is the rate constant for protein synthesis; will vary depending on the particular protein C is the amount of Protein at any time KD is the first order rate constant of enzyme degradation, i.e., the fraction destroyed per unit time, also depends on the particular protein
dC = 0 dt Steady-state is achieved when the amount of protein synthesized per unit time equals the amount being destroyed 0.693 KDC = KS t 1/2 = KD C Protein concentration (enzyme activity) Stop protein synthesis, measure rate of decay Hours after stopping synthesis
Steps in Protein Degradation ATP AMP + PPi Transformation to a degradable form (Metal oxidized, Ubiquination, N-terminal residues, PEST sequences) Lysosomal Digestion 26S Proteasome digestion 7 type, 7 type subunits Proteolysis to peptides KFERQ 8 residue fragments Ubiquination N-end rule: DRLKF: 2-3 min AGMSV: > 20 hr PEST: Rapid degradation
Glycine at C terminal of Ubiquitin Ubiquitin COO- Ubiquitin activating enzyme ATP E1 HS Ubiquitin conjugating enzyme 20 or more per cell AMP + PPi O C S E1 3 E2 SH NH3+ N HS E2 HS E1 NH3+ N H3N+ O 3 C S E2 N O O O O ATP C C C C NH Ubiquitin- specific proteases (26S proteasome) E3 AMP + PPi Poly Ubiquitin Degraded protein + Ubiquitin Activation of Ubiquitin Ubiquitin ligase Ubiquination Page 211
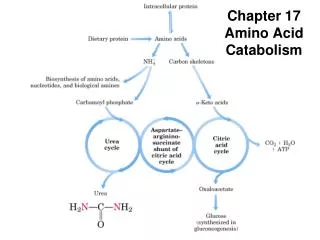
Amino Acid Catabolism Deamination of Amino Acids removal of the a-amino acids Oxidative Deamination Non-oxidative Deamination Transamination
Oxidative Deamination Only a few amino acids can be deaminated directly. Glutamate Dehydrogenase catalyzes a major reaction that effects net removal of N from the amino acid pool . Glutamate Dehydrogenase is one of the few enzymes that can utilize either NAD+ or NADP+ as electron acceptor. Oxidation at the a-carbon is followed by hydrolysis, releasing NH4+.
At right is summarized the role of transaminases in funneling amino N to glutamate, which is deaminated via Glutamate Dehydrogenase, producing NH4+.
Non-oxidative Deamination Serine Dehydratase catalyzes: serine àpyruvate + NH4
Transamination Transaminase enzymes (aminotransferases) catalyze the reversible transfer of an amino group between two a-keto acids.
Example of a transaminase reaction is shown at right. • Aspartate donates its amino group, becoming the a-keto acid oxaloacetate. • a-Ketoglutarate accepts the amino group, becoming the amino acid glutamate. An
In another example shown at right, alanine becomes pyruvate as the amino group is transferred to a-ketoglutarate.
Transaminases equilibrate amino groupsamong available a-keto acids. This permits synthesis of non-essential amino acids, using amino groups derived from other amino acids and carbon skeletons synthesized in the cell. Thus a balance of different amino acids is maintained, as proteins of varied amino acid contents are synthesized.
The The prosthetic group of the transaminase enzyme is pyridoxal phosphate (PLP), a derivative of vitamin B6. Mechanism of Transamination
In the "resting" state, the aldehyde group of pyridoxal phosphate is in a Schiff base linkage to the e-amino group of an enzymelysine residue.
The a-amino group of a substrate amino acid displaces the enzyme lysine, to form a Schiff base linkage to PLP. • The active site lysine extracts a proton, promoting tautomerization (shift of the double bond), followed by reprotonation with hydrolysis.
What was an amino acid leaves as an a-keto acid. The amino group remains on what is now pyridoxamine Phosphate (PMP). • A different a-keto acid reacts with PMP, and the process reverses, to complete the reaction.
Purine Nucleotide Cycle The activity of L-glutamate dehydrogenase is low in the skeletal muscle and heart. In this tissues purine nucleotide cycle Figure 9-7 page 216
Metabolism of One Carbon Units One carbon units are one carbon containing groups produced in catabolism of some amino acids. Methyl (-CH3), methylene (=CH2), formyl (O=CH-) and formimino (HN=CH-)
tetrahydrofolate (FH4) One carbon units are carried by tetrahydrofolate (FH4), a reduced form of folic acid.
tetrahydrofolate (FH4) • FH4 is formed in reduction of folic acid catalyzed by dihydrofolate reductase. The four hydrogens are added to the four atoms of folic acid in positions 5 to 8. The N5 and N10 nitrogen atoms of FH4 participate in the transfer of one carbon groups
Production of One Carbon Units Either glycine or serine can act as methylene donor, giving N5,N10-methyleneTHF. This behaves as "virtual formaldehyde" H2C=O in reactions. The oxidation level can be changed to methyl or methenyl by reduction or oxidation; methenylTHF can be hydrolyzed to formylTHF.
Production of One Carbon Units from Histidine • N5-formimino-tetrahydrofolate, produced in the pathway for degradation of histidine
In the pathway of histidine degradation, conversion of N-formiminoglutamate to glutamate involves transfer of the formimino group to tetrahydrofolate (THF), yielding N5-formimino-THF.
Adenosylmethionine (SAM) • S-adenosylmethionin (SAM) is the major donor of methyl group. FH4 can carry a methyl group on its N5 atom, but its transfer potential is too low for most biosynthetic methylation. • The activated methyl donor is SAM, which is synthesized by the transfer of an adenosyl group from ATP to the sulfer atom of methionine. The S-adenosylhomocysteine is formed when the methyl group of SAM is transferred to an acceptor.
Metabolism of Methionine, Cysteine and Cystine Sulfur-containing amino acids Methionine is an essential amino acid
Methionine cycle and methylation • In methionine cycle, the adenosyl group of ATP is transferred to a sulfur atom of methionine by methionine adenosyltransferase to form S-adenosylmethionine (Sam)
Methionine cycle and methylation All phosphates of ATP are lost in this reaction. The sulfonium ion of methionine is highly reactive and the methyl group of SAM is good leaving group. SAM then transfers the methyl group to some acceptors for their methylation by methyltransferase.