Download

1 / 24
270 likes | 481 Views
The unfolded protein response signals through high-order assembly of Ire1. Alexei V. Korennykh 1 , 3 , Pascal F. Egea 1 , Andrei A. Korostelev 4 , Janet Finer-Moore 1 , Chao Zhang 2 , 3 , Kevan M. Shokat 2 , 3 , Robert M. Stroud 1 & Peter Walter 1 , 3 Alysia Birkholz 1.15.2010.

E N D
The unfolded protein response signals through high-order assembly of Ire1 Alexei V. Korennykh1,3, Pascal F. Egea1, Andrei A. Korostelev4, Janet Finer-Moore1, Chao Zhang2,3, Kevan M. Shokat2,3, Robert M. Stroud1 & Peter Walter1,3 Alysia Birkholz 1.15.2010
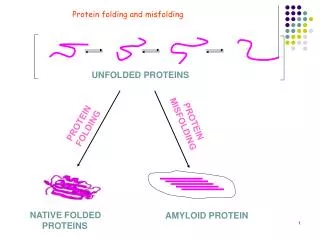
~33% of proteins fold in the ER • Need a mechanism to detect misfolded proteins • UPR • Ire1 is the crucial factor • Has been shown to dimerize • Organized into UPR induced foci • RNase • Trans-autophosphorylation • Binding of ADP
Ire1 “This pathway is really a sort of duckbilled platypus. There have been so many surprises.” - Feroz Papa UCSF • Inositol-requiring protein-1 • ER-membrane-resident kinase/RNase • a non-spliceosomal mRNA splicing reaction • In yeast HAC1 mRNA • HAC1p proteins are then translated • In metazoans, Xbp1 mRNA • Xbp1 proteins are then translated • These proteins adjust protein folding
In stressed cells, IRE-1 binds to unfolded proteins, dimerizes and trans-autophosphorylates in kinase domain This activates the RNase domain which splices XBP1 or HAC1 The spliced form of these genes leads to activation of the UPR BIOC/MCB 568 -- University of Arizona
Activation of Ire1 by self-association. Oligomerization has been suggested to promote the activation the kinase and RNase domains Binding sites of Ire1 in metazoans (right) and yeast (left)
Aim • Authors didn’t like the proposed model that had the activation loops too far away for trans-autophosphorylation • Previous model had no linker component • A different Ire1 dimer is needed for the trans-autophosphorylation
RNase activity assay: Construct cleaved is labeled with P32 and is derived from Xbp1 mRNA
Linker controls the oligomerization Oligomerization is inhibited by increasing salt concentrations- repressing RNase activity
Structure of the oligomer • Difficult to crystalize • ADP association wasn’t working • Tried using kinase inhibitors – insert into ADP binding site Kinase inhibitors actually activated the RNase activity
Initially got a Ire1- APY29 complex of 3.9 A • When they deleted 28 amino acids forming a loop, got a resolution of 3.2 A
Electron density map for APY29 bound to IRE1KR32Δ28 3 possible hydrogen bonds formed Although APY29 fits in the ADP site reasonably well, it doesn’t have the divalent Mg 2+ RNase activity of APY29 doesn’t change with addition of EDTA
14 Ire molecules formed a crystal lattice with back to back dimers
Why is this model attractive? • Kinasedimers are side by side, making trans-autophosphorylation more logical • Kinase inactivating mutant caused no phosphorylation to occur, indicating phosphorylation activity is regulated through self • Phosphorylation sites are on activation loops which face IF3c • IF3c proposed to transfer phosphates
Do all three interfaces control RNase activity? Four monomers combined Contacts within the different interfaces Single monomer
The mechanism • Initially showed that the IreKR32 seemed to show preference for double stem loop mRNA • To look at this, generated a 354 ntd RNA with a single stem loop, and a 58 ntd RNA with a double stem loop
The mechanism Ire1 has no preference for single or double loops
Localization of Helix loop helix near IF2 interface (dimerization interface) – may control RNase activity Cavity is created that is characteristic for substrate binding pockets of enzymes
Conclusions • Old model is incomplete • Ire1 kinase, Rnase, and partial linker forms a supermolecular structure • Co-factor binding and phosphorylation enhance self-association of Ire1 but neither is required • Linkers are necessary for oligomerization
Future work • Although the authors used an increased structure for crystallization, we are still missing the full structure • Need to have evidence that these supermolecular structures are actually present in in vitro/ in vivo models ?