Download

1 / 43
440 likes | 563 Views
Topics. RNA processing and regulation Network interactions in gene expression RASL-DASL the genome: approaches for systems biology. rRNA Processing Pathway: Involvement of A Series of Endo- and Exonucleases. 18S. 5S. 28S. 45S. 41S. 32S. 20S. 5S. 28S. 18S. Maturation of tRNAs.

E N D
Topics RNA processing and regulation Network interactions in gene expression RASL-DASL the genome: approaches for systems biology
rRNA Processing Pathway: Involvement of A Series of Endo- and Exonucleases 18S 5S 28S 45S 41S 32S 20S 5S 28S 18S
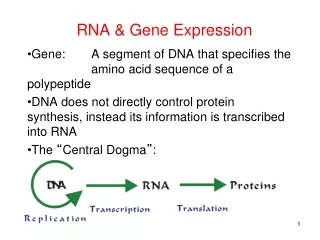
Maturation of tRNAs • tRNA is transcribed by RNA Pol III. • The 5’ end is generated by RNase P, an RNA enzyme. • The 3’ end is generated by RNase D followed by post- • transcriptional addition of CCA. • Many nucleotides are modified, which are critical for • matured tRNAs to function in translation. • Some tRNA genes contain a single intron, which is removed by • endonuclease and RNA ligase.
Transcription Unit in Eukaryotes Transcription Polyadenylation Exon (100 - 300 nts) Promoter 5’ 3’ 5’ 3’ Intron (103 - 104 nts)
pppNpNp Phosphatase Pi ppNpNp GTP Guanyl transferase PPi Gp ppNpNp Transfer of methyl group to the cap Methylase ppNpNp CH3-Gp CBP80/20 Addition of a Cap to the 5’ End of Transcript
Cleavage & polyadenylation specificity factor (CPSF) Cleavage stimulation factor (CstF) 73K 100K 50K 160K PAP 77K 30K GU-rich AAUAAA Cap 64K Cleavage site Polyadenylation signals Keller and Minvielle-Sebastia, Curr. Opion, Cell Biol. 9:329-336, 1997
The Polyadenylation Pathway • PAP stimulates cleavage by CPSF • Bound PAP adds A residues at a slow • rate to the 3'OH group • Binding of poly(A) binding protein II • accelerates A addition • PBBII plays a role in signaling poly(A) • of about 200-250 A residues From Mole. Cell Biol., Lodish et al., 2000
R: Purine Y: Pyrimidine N: Any base Consensus Splicing Signals polypyrimidine tract exonic enhancer exonic silencer 5’ splice site branchpoint 3’ splice site A/CAG GURAGU YNYURAY--Y10-20--YAG exon intron exon
Pre-mRNA A 2’ HO P P Step I: 5’ splice site cleavage and branch formation pA pA Lariat intermediate P 3’ OH Step II: 3’ splice site cleavage and exon ligation P 3’ HO + Ligated exons Released lariat intron The Nuclear pre-mRNA Splicing Pathway
U1 U1 U2 A U6 U5 U1 U4 U2 U6 U6 U5 U5 U2 U2 The Spliceosome Assembly Pathway E (Commitment Complex) Exon 1 Exon 2 ATP A (Pre-spliceosome) B (spliceosome) U4 C (Activated Spliceosome) Nuclear Export Exon 1 Exon 2 mRNA
Mechanistic Similarity between Splicing of Nuclear pre-mRNAs and Self-splicing of Group II Introns
Establishment of the Catalytic Core in the Spliceosome and Similarity to the Catalytic Core in Group II Introns Nilsen, T.W. In RNA Structure and function, 1998
Alternative Splicing: A Genomic Issue % of alternatively spliced genes # of isoforms per gene Human: 59% 3.17 Worm: 22% 1.34 Source: Nature 409:898, 2001
5'ss 3'ss 1. Balanced Splicing: 5'ss 5'ss 3'ss 2. Alt. 5' Splicing: 5'ss 3'ss 3'ss 3. Alt. 3' Splicing: 5'ss 3'ss 5'ss 3'ss 4. Exon Skipping: 5'ss 3'ss 5. Mutual Exclusion: Different Types of Alternative Splicing
Examples of Biologically Important Alternative Splicing Events Smith and Valcarcel, TIBS 25:381-388, 2000
Dscam: An Exon Guidance Receptor with 38,016 Isoforms Generated by Alternative Splicing Exon 4 Exon 9 Exon 17 Exon 6 1 12 1 48 1 33 1 2 TM Genomic DNA Protein Schmucker. D. et al., Cell 101:671-684, 2000
+ - Sxl stop stop + Sxl Tra/Tra-2 SR Dsx Dsx + Drosophila Sex Determination Pathway X:A ratio: 2:2 1:2 Sxl Sxl off tra (truncated) Tra Tra-2 Negative regulator of male differentiation genes Negative regulator of female differentiation genes
KH-type KSRP, Nova-1, PS1 KH No CELF Factors CUG-BP1, 2, ETR-3, NAPOR RRM No Four Classes of RNA Binding Proteins implicated in Splicing Regulation Family Name Examples Key Domain Required for Splicing SR Proteins SC35, ASF/SF2, 9G8, hTra2-a, hTra2-b, SRp20, 30c, 40, 46, 54, SRp55, 75, 86 RRM and RS Yes RRM, some with RGG boxes HnRNPs hnRNP A/B, hnRNP F, H hnRNP I/PTB, nPTB No
SF2/ASF hnRNP or A/B U2AF SC35 3' splice site ESE ESS Positive and Negative Influences of Splice Site Selection by Exonic Enhancers and Silencers
Dis. 5'ss Prox. 5'ss 3'ss S100 + SC35 S100 + hnRNP A1 hnRNP A1 SC35 _ _ NE M Pre-mRNA Prox. mRNAs Dist. 1 2 3 4 5 6 7 8 9 10 11 12 SR and hnRNP proteins affect alternative splicing in opposite ways Fu, X-D., et al., PNAS 89:11224-11228, 1992
Tissue-specific Alternative Splicing of the src pre-mRNA: Blockage of Splice Sites by PTB Chou, M-Y., et al., Mole. Cell 5:949-957, 2000
FGFR2 FGF FGFR2 Fibroblasts other cells Epithelial Cells 7 8 9 10 KGFR (or FGFR7) KGF KGFR Cell-specific Alternative Splicing of the FGFR2 Gene Establish an Autocrine Loop Critical for Development
CIC-1 U/G(n) Stop 2 6b 7a 7 8 3 6 Myotonic Dystrophy: A Splicing Disease Phenotype: Skeletal muscle hyperexcitability and progressive muscle wasting Cause: CUG or CCUG expansion in 3' untranslated regions in DMPK or ZNF9 genes Mechanisms: Defects in splicing of the muscle-specific chloride channel CIC-1 How? DMPK CUG-BP1 (increased stability and nuclear localization) AAAAA CUG(n) Nonsense-mediated mRNA decay & protein truncation Reduced Cl conductance Membrane hyperexcitability Mankodi, A., et al., Mole. Cell 10:35-44, 2002 Charlet-B, N., et al., Mole. Cell 10:45-53, 2002
Life, Sex, and WT1 Isoforms: Three Amino Acids Can Make All the Difference (Hastie, Cell 106, 391, 2001) All cells express +KTS and -KTS isoforms; Double heterozygous mice are normal! { +/- mice develop Frasier syndrome -/- mice die after birth with kidney defects Function as a transcriptional factor Complete male-to-female reversal Reduced Y-specific Sry expression - KTS c Exon 9 Exon 10 CATACAG GTAAAACAA gtgcgtaaactt K T S C G { +/- mice are normal -/- mice die after birth with kidney defects Function in pre-mRNA processing Undifferentiated gonad + KTS Hammes, A., et al., Cell 106:319-329, 2001
Topics RNA processing and regulation Network interactions in gene expression RASL-DASL the genome: approaches for systems biology
Complex Network of Coupled Interactions in Gene Expression T. Maniatis & R. Reed, Nature 416:499-506, 2002
Electron Microscopic Analysis of Chromosomal Spreads: Evidence for Co-transcriptional Splicing Beyer and Osheim, Genes Dev. 2:754-765, 1988
Targeting of Splicing Factors to Nascent Transcripts Depends on Pol II CTD Cell lines were constructed to express wt and CTD-truncated a-amanitin-resistant Pol II Endogenous Pol II was inhibited by a-amanitin Nascent transcripts (pem) was detected by in situ hybridization Splicing factors were localized by using specific antibodies Results show that CTD is required for the recruitment of splicing factors to the site of transcription Misteli and Spector, Mole. Cell 3:679-705, 1999
Hyperhosphorylated Pol II (IIo) Stimulates Pre-mRNA Splicing in vitro Pol II by itself has no splicing activity Hyperphosphorylated pol II (Iio), but not IIA, stimulates splicing in vitro Pol II by itself has no splicing activity (b, lanes 1, 5). CTD is necessary, but not sufficient for the effect (c, lanes 5-8, 12-25). Hirose et al., Genes Dev. 13:1234-1239, 1999
Promoter-dependent Alternative Splicing Cramer et al., Mole. Cell 4: 251-258, 1999
Splicing-derived mRNAs are More Efficiently Exported A. Spliced mRNA is exported more efficiently in injected Xenopus oocytes. B. Isolation of RNP complex containing spliced mRNA or a mRNA assembled in extracts. C. Purified mRNP is more efficiently exported than assembled mRNA-protein complexes. Luo and Reed, PNAS 96:14937-14942, 1999
RNA Metabolism in the Nucleus: Coupling RNA Splicing to Nuclear Export Nucleus Normal pre-mRNA Partial or Mutant pre-mRNA hnRNPs SR SR SR Retained and degraded Spliced mRNP introns SR SR SR Aly Aly Export complex TAP hGle2 p15 Cytoplasm Retained and degraded hDbp5 hGle Exported Reed and Magni, Nature Cell Biol 3:E201-204, 2001
eIF4F (= eIF4E, eIF4A, and eIF4G) PABP AAAAAA m7Gppp 5’UTR AUG UAA 3’UTR Circular mRNA in vivo Implications: PolyA binding protein is required for efficient translation RNA decay in many cases are translation-dependent Deadenylation will result in decapping Tharun and Parker, 1997
Initiation ATG 5' UTR CAP 4E 4G Elongation Recycling PABP PABP PABP eRF3 AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA eRF1 STOP 3' UTR Model for translation stimulation by 5'-3' interactions
m7G m7G UAA UAA UAA UAA UAA UAA UAA AUG AUG AUG AUG AUG AUG AUG AAAAAAAAAlong AAAAAAAAAlong AAAAAAAAAlong m7G AAAoligo m7G AAAoligo AAAoligo mRNA decay pathways in eukaryotic cells Deadenylation-indep. decapping Endonucleolytic Cleavage (Rnase L ?) m7G PolyA shorting (PAN) UAA Decapping 3’ to 5’ decay Decapping (Dcp1p) 3’ to 5’ decay m7G 5’ to 3’ decay 5’ to 3’ decay (Xrn1p) Tharun and Parker, 1997
- Dcp1p + Mrt1p Mrt3p Pab1p m7Gppp UAA AAAAAAlong AUG Dcp1p + Upf2 Upf1 Upf3 UAA AAAAAAlong AUG UAA m7Gppp Decapping Mechanisms A.Positive and negative effect on the decapping enzyme B.Nonsense-mediated recruitment of the decapping enzyme Tharun and Parker, 1997
3’- 5’ exoriboucleases Rep42 Rrp4 Rrp41 Rrp45 Rrp40 Rrp43 KSRP TTP Csl4 Rrp44 Mtr3 Rrp46 m7Gppp AAAAAA ARE The Exosome and its Target Chen, C-Y., et al., Cell in press 2001 3’-5’ degradation van Hoof and Parker, Cell 99:347-350, 1999
Nonsense-mediated RNA Decay: Where does It Occur? Stops at position 39, 60-61, but not at 101 and 141, render mRNAs unstable. NMD is detectable in both the nucleus and the cytoplasm Zhang, J. et al., RNA 4:801-815, 1988
Nonsense-mediated RNA Decay: The 50-55 nt Rule Zhang, J. et al., RNA 4:801-815, 1988
Stop Exon-exon junction D: > 50-55 nts: NMD < 50-55 nts: no effect Exon-exon junction D = 20-24 nts Two Positional Rules The 50-55 nts Rule for NMD The 20-24 nts Rule for Post-splicing Marker
Exon-exon Junction Complex that Links Splicing, Export, and NMD Kim, V.N. et al., Science 293:1832-1836, 2001
Summary All RNAs (rRNA, tRNA, and mRNA) are matured in a series of processing steps after transcription. mRNA processing takes place in the spliceosome, a large step-wise assembled ribonucleoprotein machinery. Alternative splicing is very common and plays an important role in development and disease. Many reactions in the nucleus are mechanistically coupled. Some proofreading mechanisms are operating to ensure the quality of processed RNAs before they are exported out of the nucleus. The EJC complex connects nuclear processing to translation and stability of mRNA in the cytoplasm.