Download
1 / 68
680 likes | 803 Views
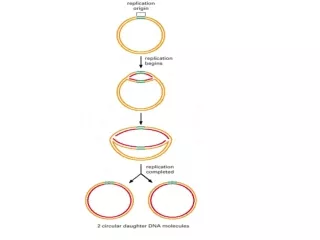
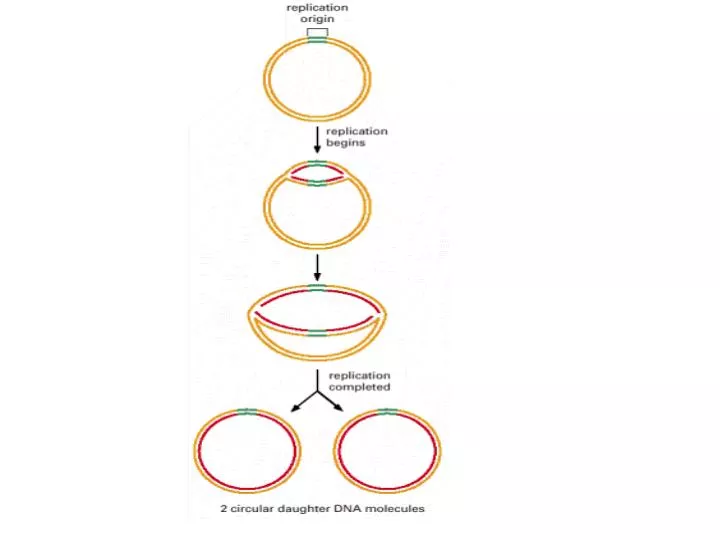
Semi consevative SSB (single-Stranded binding protein) Bidirectional Sliding rings Origin of Replication (Ori C) Clamp, clamp loader Growth fork

E N D
Semi consevative SSB (single-Stranded binding protein) Bidirectional Sliding rings Origin of Replication (Ori C) Clamp, clamp loader Growth fork Dna A, Dna B (Hexamer) Dna C Leading Strand Lagging Strand Okazaki Fragments DNA Poly I, II, III 5’----3’ Polymrase 3’----5’ Exonuclease 5’----3’ Exonuclease Klenow Fragment Proof reading RNA Primer Primase Primosome (Primase+ Helicase) DNA Ligase
Histone DNA Polymerase α,β,γ, δ, ε) ORC (Origin recognition complex) ARS (Autonomously Replicating Sequence) Core element (11 bp) PCNA (clamp) RFC ( Clamp loader) RNase H, Fen 1
Model of initiation of replication at E. coli oriC. The 9-mers and 13-mers are the repetitive sequences. Multiple copies of DnaA protein bind to the 9-mers at the origin and then “melt” (separate the strands of) the 13-mer segments. The sole function of DnaC is to deliver DnaB, which is composed of six identical subunits, to the template. One DnaB hexamer clamps around each single strand of DNA at oriC, forming the prepriming complex. DnaB is a helicase, and the two molecules then proceed to unwind the DNA in opposite directions away from the origin
OriC has a length of 245 bp. It contains a tandem array of three nearly identical 13-nucleotide sequences (green) and four binding sites (yellow) for the dnaA protein. The relative orientations of the four dnaA sites are denoted by arrows.
Consensus sequence of the minimal bacterial replication origin based on analyses of genomes from six bacterial species. Similar sequences constitute each origin; the 13-bp repetitive sequences (orange) are rich in adenine and thymine residues. The 9-bp repetitive sequences (brown) exist in both orientations; that is, the lower-right sequence, read right to left, is the same as that of the upper-left sequence, read left to right. These sequences are referred to as 13-mers and 9-mers, respectively. Indicated nucleotide position numbers are arbitrary
DNA is not synthesised in the 3′ to 5′ direction, as erroneously shown on the left-hand strand
Helicase Structure.The bacterial helicase PcrA comprises four domains: A1, A2, B1, and B2. The A1 domain includes a P-loop NTPase fold, whereas the B1 domain has a similar overall structure but lacks a P-loop and does not bind nucleotides. Single-stranded DNA binds to the A1 and B1 domains near the interfaces with domains A2 and B2.
Two metal ions (typically, Mg2+) participate in the DNA polymerase reaction. One metal ion coordinates the 3′-hydroxyl group of the primer, whereas the phosphate group of the nucleoside triphosphate bridges between the two metal ions. The hydroxyl group of the primer attacks the phosphate group to form a new O-P bond
DNA polymerases donate two hydrogen bonds to base pairs in the minor groove. Hydrogen-bond acceptors are present in these two positions for all Watson-Crick base pairs including the A-T base pair shown.
Proofreading. The growing polynucleotide chain occasionally leaves the polymerase site of DNA polymerase I and migrates to the exonuclease site. There, the last nucleotide added is removed by hydrolysis. Because mismatched bases are more likely to leave the polymerase site, this process serves to proofread the sequence of the DNA being synthesized.
Core Enzyme: subunit: DNA polymerase ε subunit:3’---5’ exonuclease θ subunit: Stimulate ε exonuclease Holoenzyme: β subunit:sliding clamp δ subunit : binds to β
Shape Selectivity. The binding of a nucleoside triphosphate (NTP) to DNA polymerase induces a conformational change, generating a tight pocket for the base pair consisting of the NTP and its partner on the template strand. Such a conformational change is possible only when the NTP corresponds to the Watson-Crick partner of the template bas
Structure of the Sliding Clamp. The dimeric β2 subunit of DNA polymerase III forms a ring that surrounds the DNA duplex. It allows the polymerase enzyme to move without falling off the DNA substrate
DNA Polymerase Structure. The first DNA polymerase structure determined was that of a fragment of E. coli DNA polymerase I called the Klenow fragment. Like other DNA polymerases, the polymerase unit resembles a right hand with fingers (blue), palm (yellow), and thumb (red). The Klenow fragment also includes an exonuclease domain
Helicase Structure. The bacterial helicase PcrA comprises four domains: A1, A2, B1, and B2. The A1 domain includes a P-loop NTPase fold, whereas the B1 domain has a similar overall structure but lacks a P-loop and does not bind nucleotides. Single-stranded DNA binds to the A1 and B1 domains near the interfaces with domains A2 and B2
Helicase Mechanism. Initially, both domains A1 and B1 of PcrA bind single-stranded DNA. On binding of ATP, the cleft between these domains closes and domain A1 slides along the DNA. On ATP hydrolysis, the cleft opens up, pulling the DNA from domain B1 toward domain A1. As this process is repeated, double-stranded DNA is unwound.
Conserved Residues among Helicases. A comparison of the amino acid sequences of hundreds of helicases revealed seven regions of strong sequence conservation (shown in color). When mapped onto the structure of PcrA, these conserved regions lie along the interface between the A1 and B1 domains and along the ATP binding surface
The images show the crystal structure of Klenow fragment (top row) and Klenow fragment complexed with DNA (bottom row) solved to 2.8 and 3.2 Å,
The DNA polymerase III holoenzyme consists of the core enzyme, the b sliding clamp and the clamp-loading complex. • the b sliding clamp The above images show different views of the beta subunit dimer. The central hole is evident in all three images. The top right-hand image shows a detailed model of the two polypeptides. The topleft -hand image shows the polypeptide backbones of the two subunits. The bottom left hand image shows the secondary structure of the polypeptides. The interior of the dimer is lined with alpha-helices; the exterior is formed by beta sheets. The bottom right hand image shows how the beta polypeptide can form a sliding clamp on DNA.
The clamp-loading complex consists of the delta (δ) (38.7 kD) and delta (δ') prime (36.9 kD), chi (χ) (16.6 kD) and psi (ψ) (15.2 kD) subunits and either or both of the gamma (γ) subunit (68.4 kD) and the tau (Τ) subunit (71.1 kD).Both the gamma (γ) subunit and the tau(Τ) subunit are 'motor' ATPases.The δ subunit binds to the β subunit and, in concert with the δ' and γsubunits and with ATP, it catalyses the opening of the βdimer to permit passage of DNA.
DNA Replication. The formation of a phosphodiester bridge is catalyzed by DNA polymerases.