Download
1 / 68
690 likes | 1.19k Views
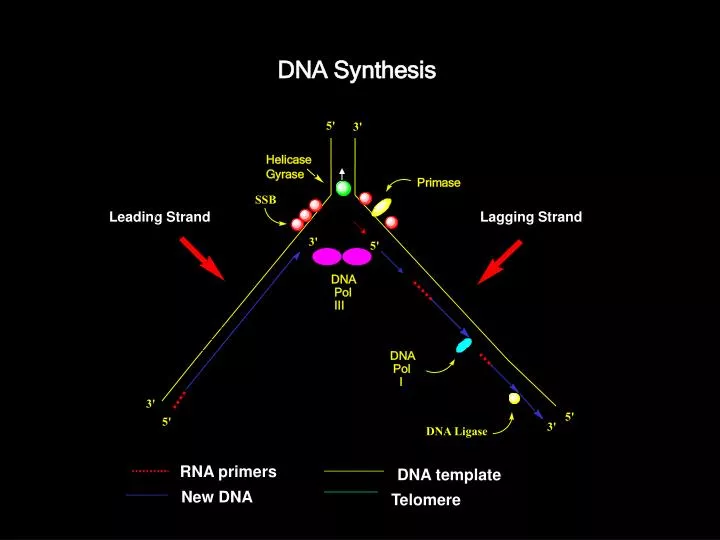
DNA Synthesis. SSB. Primase. DNA . Pol . I. DNA Ligase. 5'. 3'. Helicase Gyrase. Leading Strand. Lagging Strand. 3'. 5'. DNA . Pol . III. 3'. 5'. 5'. 3'. RNA primers. DNA template. New DNA. Telomere . E. coli DNA Polymerase III. Processive DNA Synthesis

E N D
DNA Synthesis SSB Primase DNA Pol I DNA Ligase 5' 3' Helicase Gyrase Leading Strand Lagging Strand 3' 5' DNA Pol III 3' 5' 5' 3' RNA primers DNA template New DNA Telomere
E. coli DNA Polymerase III • Processive DNA Synthesis • The bulk of DNA synthesis in E. coli is carried out by the DNA polymerase III holoenzyme. • • Extremely high processivity: once it combines with the DNA and starts polymerization, it does not come off until finished. • • Tremendous catalytic potential: up to 2000 nucleotides/sec. • • Low error rate (high fidelity) 1 error per 10,000,000 nucleotides • Complex composition (10 types of subunits) and large size (900 kd)
Sliding clamp clamp loader Polymerase Polymerase • 3'-5' exonuclease E. coli Pol III: an asymmetrical dimer Stryer Fig. 27.30
DNA Synthesis SSB Primase DNA Pol I DNA Ligase 5' 3' Helicase Gyrase Leading Strand Lagging Strand 3' 5' DNA Pol III 3' 5' 5' 3' RNA primers DNA template New DNA Telomere
Lagging strand loops to enable the simultaneous replication of both DNA strands by dimeric DNA Pol III Stryer Fig. 27.33
DNA Ligase seals the nicks O -O P O DNA Ligase + O- (ATP or NAD+) AMP + PPi O O P O OH O- • Forms phosphodiester bonds between 3’ OH and 5’ phosphate • Requires double-stranded DNA • Activates 5’phosphate to nucleophilic attack by trans-esterification with activated AMP
ENZYME O (+)H N 2 Ade P O O(-) 2.E-AMP + P-5’-DNA O AMP-O 5'-DNA P O O O O- 5'-DNA P O AMP-O OH OH O- DNA Ligase -mechanism • E + ATP E-AMP + PPi O 3. DNA-3' OH DNA-3' 5'-DNA + O P O O- + AMP-OH
DNA Synthesis in bacteria: Take Home Message 1) DNA synthesis is carried out by DNA polymerases with high fidelity. 2) DNA synthesis is characterized by initiation, priming, and processive synthesis steps and proceeds in the 5’ 3’ direction. 3) Both strands are synthesized simultaneously by the multisubunit polymerase enzyme (Pol III). One strand is made continuously (leading strand), while the other one is made in fragments (lagging strand). 4) Pol I removes the RNA primers and fills the resulting gaps, and the nicks are sealed by DNA ligase
DNA replication in eukaryotes • Similarities with E.coli replication • Polynucleotide chains are made in the 5’ 3’ direction • Require a primer (RNA). • Similarities with the E Coli DNA Pol active site and tertiary structure • Differences • Eukaryotic replication is much slower (only 100 nt/sec). • Many replication origins. • DNA is associated with histones. • DNA Polymerases are more specialized, and their interactions • are more complex. • Chromosomal DNA is linear -> requires special processing of • the ends.
Analogy between bacterial and eukaryotic proteins involved in DNA replication Bacteria Eukaryotes SSB RPA Pol I polymerase Pol a Pol III polymerase Pol d b2 subunit of Pol III PCNA 3’ exonuclease of Pol I RnaseH + FEN1 g subunit of Pol III RCF RPA = Replication protein A PCNA = proliferating cell nuclear antigen FEN1 = flap endonuclease
Lagging strand synthesis in eukaryotes RNA primer 5’ (a) RPA=Replication protein A RPA Pola/primase 10-30 nt (b) 5’ PCNA RCF RCF = clamp loader PCNA = sliding clamp (c) 5’ Pold 5’ (d) Rnase H/FEN1 RnaseH = 5’-nuclease FEN1 = flap endonuclease (e) ligase (f)
5‘… 3' 3‘… 5' Telomerase preserves chromosomal ends • The ends of the linear DNA strand cannot be replicated due to the lack of a primer • This would lead to shortening of DNA strands after replication RNA primer • Solution: the chromosomal ends are extended by DNA telomerase This enzyme adds hundreds of tandem repeats of a hexanucleotide (AGGGTT in humans) to the parental strand: 5‘… AGGGTTAGGGTTAGGGTT… 3' 3‘… 5' telomere 5‘… AGGGTTAGGGTTAGGGTT… 3' 3‘… 5' TCCCAATCCCAATCCCAA…
5‘… 3' 3‘… 5' Circular DNA does not have ends: Upstream Okazaki fragment RNA primer Linear DNA: RNA primer
Telomerase is a reverse transcriptase that uses it own RNA as a template for elongation of the 3’ end of DNA
Telomerase inhibitors • Telomerase RNA as a target for antisense drugs • G-tetraplexes at chromosomal ends as a drug target. Modified oligonucleotides that hybridize with telomerase RNA, preventing it from being used as a template for telomere synthesis. Porphyrins, anthraquinones: stabilize G-tetraplex structure, inhibit telomerase activity.
Termination of Polymerization:The Key to Nucleoside Drugs Antiviral Antitumor Principle of action: 1) cellular uptake 2) activation to 5’-triphosphate 3) incorporation in DNA resulting in chain termination
Nucleoside inhibitors of reverse transcriptase DNA RNA Proteins Cellular Action replication transcription translation DNA Typical flow of genetic information: Notable exception: retroviruses RNA RNA DNA Proteins Cellular Action Reverse transcription transcription translation RNA
Reverse transcriptases (RT) are RNA-directed DNA Polymerases Used by RNA viruses (HIV-I , human immunoblastosis virus, Rous sarcoma virus) 1. Make RNA-DNA hybrid (use its own RNA as a primer) 2. Make ss DNA by exoribonuclease (RNase H) activity 3. Make ds DNA incorporate in the host genome RT RT RT RNAse H ss DNA ds DNA RNA RNA:DNA hybrid
HIV Life Cycle 1 = Entry in CD4+ lymphocytes 2 = Reverse transcription 3 = Integration 4 = Transcription 5 = Translation 6 = Viral Assembly
Termination of Polymerization:Nucleoside Drugs (abacavir) (zidovudine) Antiviral Antitumor Other examples: dideoxycytidine, dideoxyinosine Principle of action: 1) cellular uptake 2) activation to 5’-triphosphate 3) competition with normal substrate and incorporation in DNA resulting in chain termination
Anti-HIV drug Ziagen was discovered at the U of M College of Pharmacy 1998 Robert Vince, Professor Department of Medicinal Chemistry
Nucleoside Drugs Must Be Converted toTriphosphates to be Part of DNA and RNA • Compete with normal substrate for RT binding • Cause chain termination
DNA Chain termination by Nucleoside Analogs Primer Strand Template Strand 2+ Mg O Base O P O O- O 3' OH O O O Base -O P O P O P O 5' O- O- O- Ziagen No 3’OH!
Mechanisms of selectivity • 1. Activated drug is recognized and incorporated in • DNA only by reverse transcriptase, not by cellular • DNA polymerases (RNA viruses). • viral polymerases usually have lower fidelity • (no proofreading) • Mammalian DNA polymerases are more accurate • 2. The drug is phosphorylated by viral kinase, not • by cellular kinases (e.g. AZT).
Mechanisms of resistance and possible solutions: • The drug cannot enter cells or is pumped out rapidly. • The drug is rapidly deaminated to inactive form or normal substrate is • overproduced. • 3. The drug is no longer recognized by kinases and is not • activated to triphosphate form. • Possible solution: • Use activated phosphate form of nucleosides (Viread) • 4. Activated drug is not incorporated in DNA by mutant reverse • transcriptase (usually HIV RT mutations at codons 184,65,69, 74, and 115). • Possible solution: • Use a mixture of several RT inhibitors (e.g. zidovudine (AZT) + • lamivudine (3TC) = Combivir®) or a mixture of different mechanisms • of action (e.g. non-nucleoside RT inhibitors, protease inhibitors).
Nucleoside inhibitors of DNA polymerase as anticancer drugs • used for treating acute myelocytic leukemia • activated to triphosphate form by cellular kinases • causes inhibition of DNA synthesis, repair, and DNA fragmentation • very toxic
DNA Damage, Mutations, and Repair See Stryer p. 768-773
DNA Mutations 1.Substitution mutations: one base pair for another, e.g. T for G • the most common form of mutation • transitions; purine to purine and pyrimidine to pyrimidine • • transversions; purine to pyrimidine or pyrimidine to purine • Frameshift mutations • Deletion of one or more base pairs • Insertion of one or more base pairs
NH 2 Spontaneous mutations due to DNA polymerase errors • Very low rate of misincorporation (1 per 108 - 1 per 1010) • Errors can occur due to the presence of minor tautomers of nucleobases. NH N N N HN O N N C Rare imino tautomer of A amino 10-4 Normal base pairing Mispairing
Consider misincorporation due to a rare tautomer of A 2nd replication A T 1st replication A(imino) C 5’ A T A(imino) T G C 3’ A T Normal replication Final result: A G transition (same as T C in the other strand)
Induced mutations result from DNA damage • Sources of DNA damage: endogenous • Deamination • 2. Depurination: 2,000 - 10,000 lesions/cell/day • 3. Oxidative stress: 10,000 lesions/cell/day • Sources of DNA damage: environmental • 1. Alkylating agents • 2. X-ray • 3. Dietary carcinogens • 4. UV light • 5. Smoking
Normal base pairing in DNA and an example of mispairing via chemically modified nucleobase A G T T G A G C
DNA oxidation Reactive oxygen species: HO•, H2O2, 1O2, LOO• • 10,000 oxidative lesions/cell/day in humans
O N H H N G A N N H N N N O Deamination N H O 2 N N N N H H y p o x a n t h i n e N N N N A O O N N N H N H X a n t h i n e N N N O N N H G 2 H N H O 2 N N H U r a c i l N O N O C C Rates increased by the presence of NO (nitric oxide)
Depurination to abasic sites 2,000 – 10,000/cell/day
UV light-induced DNA Damage …CC… Pyrimidine dimer Easily bypassed by Pol (eta) in an error-free manner
Deletions and insertions can be caused by intercalating agents Stryer Fig. 27.44
Metabolic activation of carcinogens N7-guanine adducts G T transversions Stryer Fig. 27.45
Chemical modifications of DNA in mutagenesis and anticancer therapy carcinogen or drug (X) detoxification metabolic activation excretion reactive metabolite (X-) DNA DNA adducts X X repair replication * * intact DNA cell death mutations Anticancer Cancer
Importance of DNA Repair • DNA is the only biological macromolecule that is repaired. All others are replaced. • More than 100 genes are required for DNA repair, even in organisms with very small genomes. • Cancer is a consequence of inadequate DNA repair.