Download
1 / 56
560 likes | 583 Views
Explore how ecological interactions shape the evolution of species through resource competition, predator-prey dynamics, and human influence, using examples like Darwin’s finches and domesticated species.

E N D
Selection in Nature • Ecological Interactions • A. Resources • - Darwin’s Finches
Selection in Nature • Ecological Interactions • A. Resources • - Darwin’s Finches
Selection in Nature • Ecological Interactions • A. Resources • - Darwin’s Finches
- Quantitative Traits like these -- affected by many genes -- have a higher probability of including a pleiotrophic gene – a gene that affects more than one trait. So, we might expect complex, quantitative traits (and their response to selection) to be CORRELATED to other traits. BUT: if selection is acting on both traits in different ways, neither will be “optimized”. Adaptations will be a compromise, depending on the relative strengths of the selective pressures, the relative values of the adaptive traits, and their heritabilities (ease with which they can respond to selection).
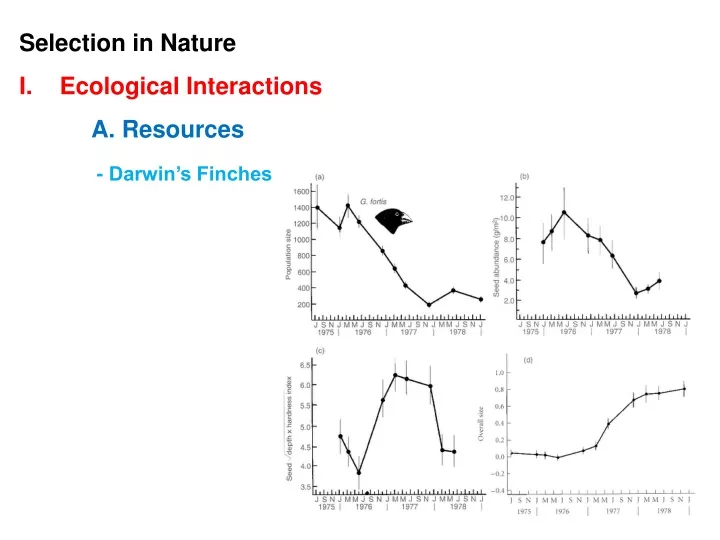
Consider the Grant’s work on medium ground finches during the drought of ‘76-’77. Birds with deep and narrow beaks had the greatest fitness.
Consider the Grant’s work on medium ground finches during the drought of ‘76-’77. Birds with deep and narrow beaks had the greatest fitness. But beak depth and beak width are POSITIVELY CORRELATED (probably developmentally).
Consider the Grant’s work on medium ground finches during the drought of ‘76-’77. Birds with deep and narrow beaks had the greatest fitness. But beak depth and beak width are POSITIVELY CORRELATED (probably developmentally). So, although selection should have pushed the pop along the blue line, it went along the green line.
Selection in Nature • Ecological Interactions • B. Competitors – “Character Displacement” • - Darwin’s Finches • (on Daphne Major)
Selection in Nature • Ecological Interactions • B. Competitors • - Darwin’s Finches Competition in sympatry leads to resource partitioning and selection for the most efficient morphology to use this subset of resources… the morphological change is “character displacement”
Selection in Nature • Ecological Interactions • B. Competitors • - Anolis (Stuart and Cambell) • (islands off east coast of FLA) Niche partitioning….
Selection in Nature • Ecological Interactions • B. Competitors • - Anolis (Stuart and Cambell) • (islands off east coast of FLA) 15 years later…patterns persisted and morphological adaptation (character displacement) had occurred Sig.
Selection in Nature • Ecological Interactions • C. Predators • - Peromyscuspolionotus • (Hoekstra) Levels of predation on clay models
Selection in Nature • Ecological Interactions • C. Predators • - Peromyscuspolionotus • (Hoekstra)
Selection in Nature • Ecological Interactions • C. Predators • - Scarlet kingsnake • (Harper and Pfennig) Stickebacks colonizing predator-free lakes lose their armor.
Selection in Nature • Ecological Interactions • C. Predators • - Scarlet kingsnake • (Harper and Pfennig) Predators only learn to avoid mimics if they experience the dangerous model.
Selection in Nature • Ecological Interactions • C. Predators • - Goldenrod gall fly • (Abrahamson)
Selection in Nature • Ecological Interactions • C. Predators • - Goldenrod gall fly • (Abrahamson) Mean gall sizes induced by flies from different families.
Selection in Nature • Ecological Interactions • C. Predators • - Goldenrod gall fly • (Abrahamson) Stabilizing selection for intermediate gall size, which is a heritable trait in both plants and flies.
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species Aurochs
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species Nagasawa et al. 2015. • Fig. 1 Comparisons of behavior and urinary oxytocin change among long gaze dogs (LG, n = 8, black bars and circles), short gaze dogs (SG, n = 22, white bars and circles), and wolves (wolf, n = 11, gray bars and square).(A) Behavior during the first 5-min interaction. (B) and (D) Changes of urinary oxytocin concentrations after a 30-min interaction. Urinary oxytocin concentrations in owners (B) and dogs or wolves (D) collected before and after a 30-min interaction are shown. (C) and (E) Comparisons of the change ratio of urinary oxytocin among LG, SG, and wolf for owners (C) and dogs or wolves (E). The results of (A), (B), and (D) are expressed as mean ± SE. (C) and (E) reflect median ± quartile. ***P < 0.001, **P < 0.01, *P < 0.05.
Fig. 2 Comparisons of behavior and urinary oxytocin between oxytocin and saline treatment conditions.(A) to (C) The effects of oxytocin administration on dog behaviors. Panels show the mean duration of dogs’ gaze at participants (A), touching participants (B), and time spent in the proximity of less than 1 m from each participant (C). Black and white bars indicate, respectively, oxytocin- and saline treatment conditions. OW, owner; UP, unfamiliar person. (D) to (G) Change in urinary oxytocin concentrations after a 30-min interaction after oxytocin or saline administration. Urinary oxytocin concentrations of owners (D) and dogs (F) before and after a 30-min interaction are shown for oxytocin and saline groups. The change ratio of urinary oxytocin in owners (E) and dogs (G) is compared between male and female dogs. ***P < 0.001, **P < 0.01, *P < 0.05. The results of (A) to (D) and (F) are expressed as mean ± SE. (E) and (G) reflect median ± quartile.
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Domesticated Species ‘selective sweep’ fixes adaptive alleles, but brings other linked alleles along for the ride – increasing homozygosity
Selection in Nature • Ecological Interactions • D. Humans as Selective Pressures • - Hunted/fished Species
Shackle et al, 2009. (a) Mean length (cm) and (b) mean mass (kg) for fish functional groups from 1970 to 2008. Linear regression equations of body size through time are shown. Dots are annual values and lines show 3-year running averages. Grey lines denote the direct measure of growth ((a) length and (b) mass at age 6 as weighted by species biomass within a functional group).
Selection for smaller fish that breed at a younger age. The problem is that small young fish produce fewer eggs.
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints Why do flying fish return to water? Gravity… and they are fish.
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints Why do larger animals have proportionally larger legs? (mass scales as the cube of length; cross-sectional area of leg, which supports the body mass, scales as the square of length. Proportional growth leads to legs to weak to support the mass.) Conforming to physics is not an adaptation.
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints Lack of genetic variation and/or maladaptive valleys:
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints • B. Contradictory Selective Pressures Photosynthetic Potential Water Availability Shady wet jungle: large leaves Sunny dry desert: Small leaves Leaf Size
Photosynthetic Potential Water Availability Shady wet jungle: large leaves Sunny dry desert: Small leaves Or none! Leaf Size
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints • B. Contradictory Selective Pressures • Survival (growth and metabolism) or reproduction? • Many small offspring or a few large ones?
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints • B. Contradictory Selective Pressures • C. Antagonistic pleiotropy • A gene has an adaptive effect for one trait or in one environment, but a deleterious effect on another trait or in another environment.
Selection in Nature • Ecological Interactions • Constraints on Selection • A. Physical / Evolutionary Constraints • B. Contradictory Selective Pressures • C. Antagonistic pleiotropy • D. Implication Not all structures are adaptations!
D. Implication Not all structures are adaptations! “The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme”” – S. J. Gould and R. C. Lewontin (1979)
D. Implication Not all structures are adaptations! “The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme”” – S. J. Gould and R. C. Lewontin (1979) “Things can not be other than they are… everything is made for their best purpose.” Dr. Pangloss, “Candide” (Voltaire)
Selection in Nature • Ecological Interactions • Constraints on Selection • III. Back to the Neutral Theory!
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions - Rates should vary in different codon positions. Variation at the third position should be higher, because these are usually silent mutations. Mutations at the second position change amino acids, and these changes are deleterious. PATTERN CONFIRMED. - Rates should vary in coding and non-coding regions. Variation in Introns should occur more rapidly than variation in exons, since introns are not transcribed and are also invisible to selection. PATTERN CONFIRMED - Rates should vary in functional and non-functional regions of proteins. PATTERN CONFIRMED
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time.
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time. - If changes are random and mutations occur at a given rate, then replacement should "tick" along like a clock. - Selection should speed rates when a new adaptive combination occurs, like in obviously adaptive morphological trait. Then inhibit further change unless it is adaptive or neutral - PATTERNS CONFIRMED (usually).
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time. 3. Rates of morphological change should be independent of the rate of molecular change.
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time. 3. Rates of morphological change should be independent of the rate of molecular change. - "Living Fossils" show extreme genetic change and variation, yet have remained morphologically unchanged for millennia. And, their rate of genetic change in this morphologically constant species is the same as in hominids, which have changed dramatically in morphology over a short period. PATTERN CONFIRMED
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time. 3. Rates of morphological change should be independent of the rate of molecular change. 4. A truly neutral clock should tick off mutations at a constant rate. But should this ticking occur per unit time, or per generation?
III. Back to The Neutral Theory A. Neutral Variation - change in protein that does not affect fitness - ‘silent’ or ‘synonymous’ mutations are the prototype B. Predictions and Results 1. Rates of molecular evolution should vary in functional and non-functional regions 2. Rates of replacement (substitution of one fixed allele by another that reaches fixation) should be constant over geologic time. 3. Rates of morphological change should be independent of the rate of molecular change. 4. A truly neutral clock should tick off mutations at a constant rate. But should this ticking occur per unit time, or per generation? - Mutations occur during DNA replication of the DNA, so a truly neutral clock should tick at a rate dependent on the generation time of the organism. Species with rapid generation times should accumulate mutations at a faster rate than long-lived species with slower generation times. - This is true of non-coding DNA... but not true for proteins. Proteins accumulate mutations in absolute time, not generational time. THIS IS INCONSISTENT WITH THE NEUTRAL MODEL
III. Back to The Neutral Theory A. Neutral Variation B. Predictions and Results C. Ohta’s “Nearly Neutral” Model