Download

1 / 30
370 likes | 773 Views
Cell biology 2014 (revised 11/2-14 ). Lecture 10 :. Cell adhesion and ECM. The eukaryotic kingdom. Cell Biology interactive media ”video” or ”animation ”. The four major tissues in the human body. Metazoan cells form organs with specialized tissues:. - Epithelial. Cells. - Muscle.

E N D
Cell biology 2014 (revised 11/2-14) Lecture 10: Cell adhesion and ECM The eukaryotic kingdom Cell Biology interactive media ”video” or ”animation”
The four major tissues in the human body Metazoan cells form organs with specialized tissues: - Epithelial Cells - Muscle - Connective Cells + ECM - Nerve
3. 3. 2. 2. 1. 1. Epidermis * Stratum Corneum Basal lamina Basal layer Dermis ECM /residual cells* (”fibroblasts”)
Different types of cell adhesion Homophilic binding Heterophilic binding
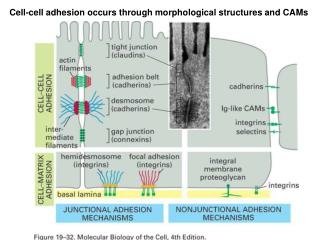
Cell-cell contacts in columnar epithelia Restricting movement of extra-cellular fluids Tight junction Adherens junction Cell-cell adhesion Desmosome Connection allowing local communication Gap junction hemidesmosome Cell-ECM adhesion Basal lamina
Glucose Glucose I. Tight junctions: function Tight junctions seal epithelial sheets to block passage of fluids in between cells Intestine Active and selective transport through the cytosol of cells by e.g., the Na+ driven glucose symport Blood vessels
II. Tight junctions: Architecture Tight junctions are made up by occludin and claudin. These are transmembrane proteins, which form tight connections across the extracellular space Linking protein attaches occludin and claudin to the cortical actin cytoskeleton The appearance of tight junctions resemble stitches across the plasma membranes of the two cells
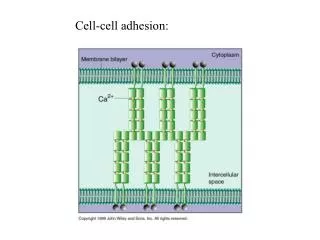
Ca2+ I. Cadherins: adherence junctions Adherence junctions from stable cell-cell adhesion points between adjacent cells a-catenin Many cadherins are known: E-cadherin in Epithelia N-cadherin in Neural cells b-catenin Cell #1 P.M. Cadherin (calcium-dependent adhesion) P.M. Cell #2 b-catenin Linkers of cadherins to the actin cytoskeleton a-catenin Actin filament video 19.1- adhesion_junctions animation 20.2-contact_inhibition
Wnt P Ub Ub Ub Cytosolic b-catenin is stabilized by Wnt signaling Frizzled LRP ZZZ Dishevelled Dishevelled GSK-3b GSK-3b Axin APC APC Axin b-catenin b-catenin Groucho Cell cycle entry + etc G1 b-catenin TCF Target genes TCF Target genes
3. 4. 2. 1. Ca2+ II. Cadherins: growth arrest at cell-cell contact Sequestering of cytosolic b-catenin at the adherence junctions formed after cell proliferation (at ”density arrest”) Wnt ..but is stabilized by Wnt signaling P.M. b-catenin enters the nucleus: G1 cyclin transcription cell proliferation b-catenin a-catenin b-catenin G1 b-catenin TCF Cytosolic b-catenin is by default unstable..... (Ubiq. dep. degradation) G1 cyclin gene
III. Cadherins: organization of cells into organs +Ca2+ Cells expressing different cadherins Cells expressing different amounts of the same cadherin +Ca2+ Cadherins are important for organ formation during development
Ca2+ Structure and function of the desmosome Desmosomes hold cells together like rivets. Through linkage to IFs, they distribute shear forces evenly within the cell Linkers Cell #1 P.M. Cadherin family protein P.M. Cell #2 Linkers Intermediate filament (IF) animation 16.4- intermediate_filament
P P P P P P Structure and regulation of gap junctions Connexon Different connexins – different pore size Connexin Cell #1 Cell #2 Regulation of pore size = ~1.5 nm Free passage of: Amino acids Nucleotides Sugars Ions ”2nd messengers”
I. Integrins: Structure and ligand specificity • Hetero-dimeric proteins consisting of a- and b- chains • ii) At least 21 cell-type specific isoforms of a/b-chain pairs • iii) Integrin ligands include ECM components (collagen, • fibronectin, laminin) and structures on neighboring cells Integrins linked to IF (hemidesmosomes: epithelia) Integrins linked to actin (focal adhesions: fibroblasts) y y(ligand: fibronectin) x x(ligand: laminin) ECM: connective tissue ECM: Basal lamina
II. Integrins: Anchorage to ECM Integrins linked to IF (hemidesmosomes) Static cell-ECM interactions , e.g. epithelial sheets ECM: Basal lamina Basal lamina: barrier towards connective tissue Integrins linked to actin (focal adhesions) Dynamic cell-ECM interactions, e.g., during migration of fibroblasts or leukocytes Inactive integrin ECM: connective tissue (contains residual migratory cells)
P P P FAK FAK III. Integrins: Architecture of the focal adhesion Focal adhesions exist only in motile cells (i.e., not in epithelia) The dynamic nature of focal adhesion is dependent on both “Inside-out” and “Outside-in” signaling FAK: Focal adhesion kinase integrin dependant signaling Linker Talin Talin P. M. Active integrin ECM Tyr- Tyr- Tyr- recruitment of SH2-domain signaling proteins (Clustering of FAK trans-phosphorylation, i.e. the same principle as for tyrosine kinase receptors, which are dimerized by ligand binding)
2. 2. 1. 1. IV. Integrins: Regulation of ligand-affinity The a- and b-chains of integrins have affinity for both “each other” and ECM ligands the concept of competing affinities Outside-in activation of ECM-binding Default state: The a- and b-chains are tightly associated Activated state: a- and b-chains are pushed apart and clustered by talin High affinity/avidity ECM-association Inside-out activation of ECM-binding
P FAK 4. 4. 2. 2. 2. 1. 1. 3. 3. 3. V. Integrins: Inside-out activation Activation of talin by a RTK ligand (e.g. EGF) Separation of a- and b-chains High affinityECM-binding Integrin clustering increased avidity Albert et al. Fig 19-49 Inactive talin • Activated talin: • Pushes a- and b-chains apart • Clusters the cytosolic parts of integrin b-chains • Links -chains with actin filaments • Recruit focal adhesions proteins (vinculin, FAK etc) • generation of a focal adhesion point
2. 2. 2. 1. 1. 3. 4. 3. 4. VI. Integrins: Outside-in activation Binding by (very) high affinity ECM ligands…….. breaks the interaction between the - and -chains The exposed -chain talin-binding site…….. ….activates talin Generation of a focal adhesion point outside-in + inside-out = positive feedback inactive talin ECM ECM ECM
P P P P P P P P P P P FAK VII. Integrins: survival and cell proliferation signals Motile cell types requires ECM for both growth and survival Survival Cell cycle entry GTP Ras Talin PI-3 K MAPK myc PKB/Akt Bad 3 -Tyr- -Tyr- -Tyr- 14-3-3 Plasma membrane
I. The extra-cellular matrix (ECM) • - Provides mechanical support to tissues • Organizes cells into tissues • ‘Instructs’ cells as to where they are and what they should do • - Reservoir for extra-cellular signaling molecules
1. 2. 3. 1. 2. 3. II. The extracellular matrix (ECM) Composed of polymeric networks of several types of macromolecules. Secreted by connective tissue cells, such as fibroblasts & chondrocytes. Proteoglycan molecules form highly hydrated gel-like “ground substance” in which the fibrous proteins are embedded Structural proteins, such as collagen and elastin, strengthen and organize the matrix Multi-adhesive proteins, such as fibronectin and laminin, facilitate cell attachment to the ECM The aqueous phase of the ECM permits diffusion of nutrients
- - - - - III. ECM: general structure of proteoglycan Linking saccharides Polysaccharide sidechain Glucosamino- glycans (GAGs) linear polymers of repeating disaccharides Protein core O-linked sugar Na+ Ca2+ Osmosis Negatively charged saccharides attract counter ions and water, giving the ECM the property to resist compression and bounce back to its original shape H2O
IV. ECM: Proteoglycan aggregates Proteoglycans can form huge aggregates onto hyaluronan. These aggregates can be up to 4 mm in length Linker protein Hyaluronan, up to 50 000 repeating disaccharides These aggregates have a very high shock absorbing capacity and are highly enriched in cartilage
V. ECM: Collagen architecture Collagen is the most common protein in body, it forms strong and flexible fibers. Many types (at least 15) Collagen a-chain (single helix) Assembled in ER Collagen molecule (triple helix) Collagen fibril Assembled outside the cell Collagen fiber
VI. ECM: Elastic elastin networks In cases there ECM is very flexible, e.g.,in skin, lungs and blood vessel walls, some of the collagen is replaced by elastin. Cross-linked elastin behaves like a rubber band! Single elastin molecule Crosslinking Stretching Relaxation
Ca10(PO4)10(OH)2 Ca10(PO4)10(OH)2 VII. ECM: different types of connective tissue ”Normal” connective tissue Bone Cartilage Osteoblast Chondrocyte Fibroblast Physical properties of the tissue depend on the content of the ECM, which is determined by the residual cell type
Summary: ECM – a sticky business! Fibronectin - Present in all ECM and primary high-affinity ligand for focal adhesions Laminin - Present in basal lamina of epithelia and the ligand for hemidesmosomes
Differential means to achieve mechanical strength Cells resistant to mechanical stress ECM (but not cells) resistent to mechanical stress Epithelial cells Basal lamina (dense ECM) Connective tissue (ECM + cells) Fig. 19-1: Epithelial tissue: The intermediate filaments of the cells themselves (linked from cell to cell by desmosomes) provides mechanical strength. Hemidesmosomes (integrin binding to laminin) are only found in the epithelial cells that connect to the basal lamina. These epithelial cells are normally essentially non-motile. Connective tissue: ECM provides the mechanical strength, the sole role of the residual cells (fibroblasts) is to produce the ECM components. These residual cells move around and may migrate to e.g. a site of tissue damage.
“Recommended reading” Chapter 19 1131-1145 1150-1162 1164-1194 Focus on the general principles and topics highlighted in the lecture synopsis Alberts et al 5th edition