Download
1 / 153
1.57k likes | 3.38k Views
熱に不安定な蛍光タンパク質を発現 39.5℃ では ER にとどまる 10℃ にすると、 folding して Exit site に集まる(温度が低いので Golgi へは移行しにくい) 20℃ では Golgi へ. Vesicle budding-fusion Vesicle flow or Membrane flow Budding → Fusion. 動物は小胞を運ぶ、 植物は Golgi を運ぶ?. Vesicle or Membrane Flow. 酵母のミュータントで詳細が解明される. ER to Golgi transport.

E N D
熱に不安定な蛍光タンパク質を発現39.5℃ではERにとどまる10℃にすると、foldingしてExit siteに集まる(温度が低いのでGolgiへは移行しにくい)20℃ではGolgiへ
Vesicle budding-fusionVesicle floworMembrane flowBudding→Fusion
ER to Golgi transport Movies: Sec_Vesicle.mov Sec_tubule_MT.mov
ERから先はかなり複雑小胞Vesicleは主に出芽の際のコートの種類で分類されるERから先はかなり複雑小胞Vesicleは主に出芽の際のコートの種類で分類される Anterograde & retrograde flows 細胞工学Vol. 25 No. 11 (2006) ARFによるメンブレントラフィックの調節
3主の小胞とそれを形成するコート (タンパク質と脂質両方を運ぶ)3主の小胞とそれを形成するコート (タンパク質と脂質両方を運ぶ)
Clathrin coat ほかの二つは寿命が短かく、特徴的な構造がないので、最近になって発見された
Cargo膜タンパクを認識・集積して形成比較的すぐに解離Cargo膜タンパクを認識・集積して形成比較的すぐに解離 MBC Figure 13-8.
Coatは3種類何が特異性を決めているか??Cargoに対してはGTPase+Adapter膜特異性では,付加されるSNARE, Tether, Rab and its effectors(>60 Rabs and >30 SNAREs)
Budding, uncoating, targeting-tethering-docking, priming & fusion Cargoの選択があるかどうかが大問題 細胞内輸送がわかる 「小胞体-ゴルジ体間輸送」
熱に不安定な蛍光タンパク質を発現39.5℃ではERにとどまる10℃にすると、foldingしてExit siteに集まる(温度が低いのでGolgiへは移行しにくい)20℃ではGolgiへ
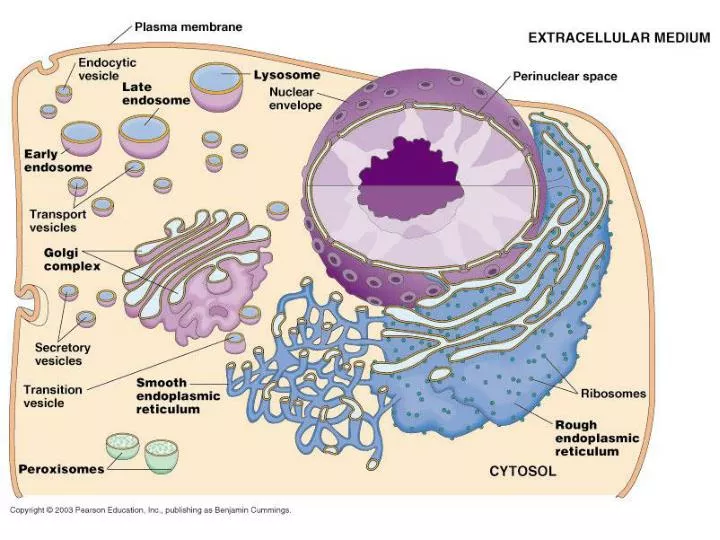
ER-Exit Site(ERES or tER)ribosome-free ER subdomains approximately 0.5 mm in diameterERGIC (ER-Golgi Intermediate compartment)orVTC (vesicular tubular cluster)Cis-Golgi ERGIC marker ERGIC-53 immunodetected. 糖鎖を認識して、可溶性のカーゴタンパクを集める! Exit siteとERGICは完全には重ならないが近接する Journal of Cell Science 119, 2173-2183 (2006)
Recruitment/selection Soluble cargo receptor: 例えばERGIC-53
Retention of incompletely folded proteins シャペロンが結合しているので、receptorに認識されない!
Exit siteで何が起こっているか 蛋白質核酸酵素 Vol. 49 No. 7 (2004) 910 Sar1 GTPase(secretion-associated and Ras-related protein) 70~90 nm vesicle Sar1-GTP→GDP SNARE, Rabなどのspecificity factorもリクルートされる
Sar1=G-proteinを足場に CargoとCoatが集合する それ以外に、小胞輸送・融合に関わる多数の因子も同時に、あるいは直後に Cargoの選別 小胞の出芽 Coatが完成(して出芽が成功)すれば、すぐにSar1は抜け落ちる?Clathrin coat以外では、Coatも比較的すぐに外れる
Sar1 Activated by Sec12 (its GEF) and anchored to membrane by its own N-terminal (above) Unhydrolyzable analogue of GTP Trends in Cell Biology13: 59-60, 2003
Activation of Sar1 by its GEF Sec12 Nature Rev Mol Cell Biol 7: 727, 2006
Prebudding complex:Sar1+Sec23/24 膜にアンカーしたSar1を起点に曲面を作る! Trends in Cell Biology13: 59-60, 2003
Volume 14, Issue 2 , April 2004, Pages 147-153 A & B Cargo-binding sites
A cargo, transport protein (red: CFTR) with diacidic motif (DXE/DXD, green) Cargoも必要!!
Outer Coat – Sec13/Sec31 b-プロペラ と a-ソレノイド Zig-zag a-helices 7: 727-738, 2006
構造がうまく組み上がると、最後には,GAP (Sec23)が働く 水 GTP analogue Sec23のR側鎖が挿入される Mg2+ Volume 14, Issue 2 , April 2004, Pages 147-153
Clathrin(アームの部分は短いゆがんだαヘリックスが作るチューブ構造)と似ているClathrin(アームの部分は短いゆがんだαヘリックスが作るチューブ構造)と似ている Coatの外層は共通の構造で、shell殻構造を作る
COPII and Clathrin nature04339-s5.mov
外殻 内層=アダプター G-protein, Cargo, 外殻との界面
Arfなし 内層=アダプター G-protein, Cargo, Coat shellとの界面
Coat GTPases: Sar1 and Arfs(ADP ribosylation factor) 外殻の構造は少しずつ異なるが 基本のモジュール構造は同じ Structural & Evolutionary aspects NCBI Books: Development
ほぼ一定のサイズの小胞ができる70~90 nm Sec23/Sec24 complex Sec13/31 complex In vitro reconstituted buds Prebudding complex on a 60 nm sphere Volume 14, Issue 2 , April 2004, Pages 147-153
Arf/Adaptor (Sec23/24) unitの数辺の数の二倍 60 nm 径(右図)で24辺、48個 90 nmとなると、下図右端で~200個 うまく数えられません! nature04339-s5.mov 64 100
いくつかの問題 tER (ERES)はどのようにできるか Cargo selectionがあるか (多分ある.でなければ,Bulk flowで間違って送られたものをCOP I小胞で戻すのみ.) But how and to what extent?? Sar1 GTP加水分解の役割(fission自体に関与?)
Sec16とCargoがExit siteを作る?Sec23-YFPで可視化 EMBO reports 4: 210-217, 2003 移動する 増える De novo formation, division & fusion
Sec12はmobile! Soderholm1.mov Pichia pastoris 蛍光は35%に Dev. Cell 6: 649-659, 2004 Fluorescent recovery after photobleaching (FRAP)
Sec12を過剰に発現させると,細胞質に(2・3段目)(saturation)他と相互作用してExit siteに集まる 発現停止後の時間 過剰 過剰
S. cerevisiaeではSec12はER全体に広がっている。PichiaのSec12を発現させても同様の挙動。( Pichiaの細胞では,それ自身でなく,他と相互作用して集まる。)逆に, S. cerevisiaeのSec12をPichiaに入れても集まらないので,構造特異性・種特異性は高い。
Regulation of Sar1 NH2 terminus by GTP binding and hydrolysis promotes membrane deformation to control COPII vesicle fission JCB, Volume 171, Number 6, 919-924 N-末端自体がリポソームに影響
(C) Sar1-GTP is capable of constricting liposome membranes. Liposomes (80–100 nm, DOPC and DLPA, 80/20 mol percent) were incubated in buffer (control; a), 10 µM Sar1-GDP (Sar1-T39N, b), 15 µM Sar1-GTP (Sar1-H79G, c), or 10 µM 9-Sar1 (d) mutants with GTP or GDP for 1 h at 37°C. (D) A gallery of Sar1-GTP–induced tubulating (a–d) and fused (e and f) DOPC/DLPA liposomes. E) Cholesterol/DOPC/DLPA (20/75/5 mol percent) liposomes (100–120 nm) were incubated as described above in the absence or presence of Sar1-GTP as indicated. (F) A gallery of tubulating cholesterol/DOPC/DLPA liposomes deformed during incubations with Sar1-GTP. Bars, 100 nm.
Sar1 Sec23 Permeabi-lized cells GTP加水分解阻害 Sec13 Sec31 GTP-gS (unhydrolyzable)
GTPase活性は膜の曲率が高くなると活性化される(別のCoat GTPase Arf1) Nature426: 507, 2003 Trp残基の蛍光:疎水環境で強い
光散乱でコート形成解離を見る 小さいリポソームほど早く解離する。と言うより turnover!