Download
1 / 67
670 likes | 792 Views
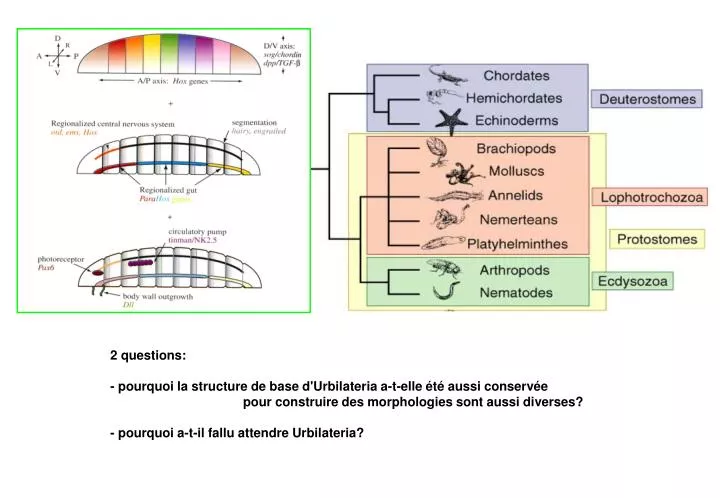
2 questions: - pourquoi la structure de base d'Urbilateria a-t-elle été aussi conservée pour construire des morphologies sont aussi diverses? - pourquoi a-t-il fallu attendre Urbilateria?. pourquoi la structure de base d'Urbilateria a-t-elle été aussi conservée

E N D
2 questions: - pourquoi la structure de base d'Urbilateria a-t-elle été aussi conservée pour construire des morphologies sont aussi diverses? - pourquoi a-t-il fallu attendre Urbilateria?
pourquoi la structure de base d'Urbilateria a-t-elle été aussi conservée pour construire des morphologies sont aussi diverses? pas seulement Urbilateria: même plus tard on observeune conservation inattendue 1. La migration de la ligne latérale 2. La ligne latérale et l'oreille 3. Le pattern des organes sensoriels d'insectes
Amphibiens (grenouille) Chondrostéens (esturgeon) Amiiformes (Amia) Elopomorphes (anguille) Clupéomorphes (hareng) Characiformes (tétra aveugle) Cypriniformes (poisson-zèbre) Salmoniformes (truite) Esociformes (brochet) Gadiformes (cabillaud) Athérinomorphes (medaka) Percomorphes (turbot)
Amphibiens (grenouille) Chondrostéens (esturgeon) Amiiformes (Amia) Elopomorphes (anguille) Clupéomorphes (hareng) Characiformes (tétra aveugle) Cypriniformes (poisson-zèbre) Salmoniformes (truite) Esociformes (brochet) Gadiformes (cabillaud) Athérinomorphes (medaka) Percomorphes (turbot)
poisson-zèbre: pattern incomplet établi rapidement (2j) amphibien: pattern complet établi lentement (10j ou plus) Autant de patterns que d'espèces?
Amphibiens (grenouille) Chondrostéens (esturgeon) Amiiformes (Amia) Elopomorphes (anguille) Clupéomorphes (hareng) Characiformes (tétra aveugle) Cypriniformes (poisson-zèbre) Salmoniformes (truite) Esociformes (brochet) Gadiformes (cabillaud) Athérinomorphes (medaka) Percomorphes (turbot)
Xenopus (crapaud) Acipenser (esturgeon)
Amphibiens (grenouille) Chondrostéens (esturgeon) Amiiformes (Amia) Elopomorphes (anguille) Clupéomorphes (hareng) Characiformes (tétra aveugle) Cypriniformes (poisson-zèbre) Salmoniformes (truite) Esociformes (brochet) Gadiformes (cabillaud) Athérinomorphes (medaka) Percomorphes (turbot)
Poisson zèbre Poisson plat (turbot)
Amphibiens (grenouille) Chondrostéens (esturgeon) Amiiformes (Amia) Elopomorphes (anguille) Clupéomorphes (hareng) Characiformes (tétra aveugle) Cypriniformes (poisson-zèbre) Salmoniformes (truite) Esociformes (brochet) Gadiformes (cabillaud) Athérinomorphes (medaka) Percomorphes (turbot) aucun changement depuis 150-200 millions d'années...
La ligne latérale et l'oreille pit neuromasts canal neuromasts surface neuromasts
la cochlée est étroitement apparentée à la ligne latérale sa fonction (audition) est complètement différente
Question de départ: pourquoi la structure de base d'Urbilateria a-t-elle été aussi conservée pour construire des morphologies aussi diverses ? Que nous a appris la génétique du développement? Le développement peut être décomposé en étapes Le programme peut être décomposé en syntagmes Trois cas de conservation extrême, alors même que - l'utilisation qui en est faite varie (organes sensoriels droso et sauterelle) - la fonction change complètement (ligne latérale et cochlée) - le pattern final est très différent (ligne latérale chez différents téléostéens)
1. Les gènes du développement sont intégrés dans un réseau complexe d’interactions et leur fonction dépend de ce réseau => contrainte fonctionnelle: tout changement sera tout d’abord testé pour sa compatibilité avec la structure génétique existante. • La seule exigence absolue est que la cohérence du programme existant ne soit pas affectée • 2. Plus il y a d’interactions, plus le nombre de degrés de liberté se réduit • les changements sont de plus en plus contraints (peu de flexibilité résiduelle) - cf ribosome • vu la complexité du programme développemental de tout organisme, • les seuls changements tolérés sont ceux qui ne changent rien • => inertie génétique • 1ère règle: règle des changements conservatoires
Règle “des changements conservatoires” (= règle de cohérence) est valide pour tous les systèmes biologiques Tout changement d’une structure biologique est testé pour sa compatibilité avec le fonctionnement de cette structure ex: dans homéobox, plus de la moitié des résidus absolument conservés sont ceux qui assurent la stabilité structurale du motif ex: nombre de protéines possibles = n20 pour n ac. aminés. Non! l'ensemble des possibles est soumis à des contraintes internes (pliabilité, stabilité, reproductibilité du reploiement) ex: cerveau: tout changement est criblé pour sa compatibilité on ajoute, on réitère, on ne touche pas à ce qui existe (ex: projection LL-auditive) ex: mental: toute idée, information nouvelle est criblée pour sa compatibilité esprit critique et recherche de cohérence (ex: contes de fées)
Inertie génétique: syntagmes très conservés des syntagmes des étapes Peu de nouveautés développementales Conservation des interactions Equivalence fonctionnelle des orthologues (Hox, Pax6, bHLH...)
Par changement de la connectivité entre syntagmes Interactions au sein d'un syntagme : nombreuses et souvent bidirectionnelles Interactions entre syntagmes: limitées simples et généralement unidirectionnelles Réitération Hétérochronie Changement spatial Modulation
. . . de nouveaux organes apparaissent en une seconde vague antéropostérieure et s’intercalent entre les neuromastes de l'embryon Réitération: cas de la ligne latérale (+ modulation: changement de polarité des neuromastes)
Changement spatial: cas de la pigmentation
* * * * *
weak wing blade enhancer weak wing vein enhancer yellow +(gene controlling pigmentation) wing spot enhancer wing spot enhancer
Très facile de soumettre un module à un nouveau contrôle: il suffit de faire apparaître un nouveau enhancer (4-8bp) variation génomique (mutations aléatoires) fait constamment de nouvelles propositions - changements ds structure protéines rarement acceptés (pcq altèrent très souvent la fonction de la protéine et/ou ses interactions) - changements ds régions non-codantes ont moins souvent des effets délétères majeurs droso: si ± 1 mutation légèrement délétère par zygote (donnée expérim.) et sachant que seulement 5-10% du génome est codant, sans doute ± 10 mut. neutres par génome avec estimations réalistes sur la taille de la population mondiale et temps génération ± 1017 nouvelles propositions en 1000 ans - même s'il n'y en a qu'une sur un million , 1011 propositions intéressantes en 1000 ans… homme: ± 5 mutations spontanées par génération 25.109 propositions nouvelles à chaque génération 1012 propositions nouvelles en 1000 ans
Toute nouvelle proposition qui est raisonnablement compatible avec tout le reste du programme • sera acceptée (sélection pour la cohérence!) • Si cette mutation rend une étape particulière un tout petit peu plus robuste • elle sera avantageuse • => accumulation de mécanismes menant au même résultat (redondance développementale)
autre exemple de changement spatial
Par nouvelle utilisation de syntagmes anciens ex: déploiement des gènes Hox dans les pattes des vertébrés
Exemple de réemploi les gènes Hox sont redéployés le long des appendices, chez les vertébrés, en gardant la règle de co-linéarité
Pas de séparation des doigts dans un mutant Hoxa13 (absence d’apoptose)
Hexadactylie: jamais six doigts - toujours cinq doigts dont un dupliqué
le nombre d’osselets et de « doigts » n’était pas fixé chez les premiers tétrapodes 5 chez tous les tétrapodes actuels, quelle que soit la forme finale de la main parce que le système Hox définit 5 identités? une fois que le programme 5-doigts est devenu très robuste (peut-être en exploitant le syntagme Hox) il peut supporter d’énormes variations morphologiques mais toujours en retenant les 5 doigts, forcément!
1. seuls les changements conservatoires sont tolérés, toute addition qui n’altère rien sera acceptée • (redondance des gènes, des mécanismes, des facteurs) • tendance à la stabilisation de tout processus existant • par accumulation d’autres processus compatibles • « canalisation » (Waddington) pression sélective majeure pour la résistance au bruit et aux fluctuations 2. une fois qu’un processus est devenu extrêmement stable (robuste) il peut supporter des variations substantielles sans collapser 2ème règle: lastabilité engendre la variété Hans Othmer et Turing: not applicable because not reliable enough - could work if nothing else, then as soon as there is a better process, takes over - but must produce same result (first rule)
Urbilateria x x x x Succession de stades pas assez stables pour varier "Evolvability" Avant l'explosion cambrienne: faune d'Ediacara (600 - 545 Myrs)
Mawsonites Dickinsonia
Parvancorina - protoarthropode??? Vaguement semblable aux trilobites… Tribrachidium (sym. ordre 3) Apparenté aux cnidaires? Aux échinodermes?
Analogie: apparition de l'écriture (Stanislas Dehaene) lecture existe depuis 5.000 ans invention à peu près simultanée, et en tout cas indépendante, des hiéroglyphes, symboles maya, écritures chinoise et cunéiforme comment est apparue cette aptitude? Explication "sciences sociales" standard: lecture est activité culturelle toute la culture est plasticité, architecture du cerveau est irrelevante Comment lit-on? Différentes logiques d'écriture utilisent-elles des mécanismes différents? • neuroimagerie : c'est toujours la même région du cerveau qui est impliquée (celle atteinte chez les alexiques: peuvent écrire très bien, mais pas lire...) • il existe un territoire spécialisé dans "la" lecture - quelle que soit la symbolique utilisée Explication "Darwinienne" standard: toute aptitude en biologie est adaptative (résultat sélection naturelle) comment la capacité à lire a-t-elle pu être sélectionnée il y a 200.000 ans?
approche expérimentale: priming d'un mot par projection subliminale du même mot (p ex raccourcit le temps pour reconnaître le mot) permet de distinguer les éléments qui sont "reconnus" (permettront le priming) 1. le temps nécessaire pour reconnaître un mot est indépendant de sa longueur (sauf pour alexiques, utilisent autres mécanismes pour déchiffer lettre à lettre) 2. analyse du mot par groupes de 2 lettres (bigrammes), et sans doute de tétragrammes => ce n'est pas la forme globale du mot qui est reconnue, mais sa décomposition multiple et parallèle en bigrammes 3. comment se fait la reconnaissance des lettres individuelles?






















