Download
1 / 1
20 likes | 253 Views
Allelopathy in an invasive shrub: A comparison of European buckthorn to other species Gena Dubois, Jenna Noble, Lisa Walters, Todd Tracy Northwestern College, Orange City, Iowa. This research was funded by a grant from the Iowa Science Foundation, administered by the Iowa Academy of Science.

E N D
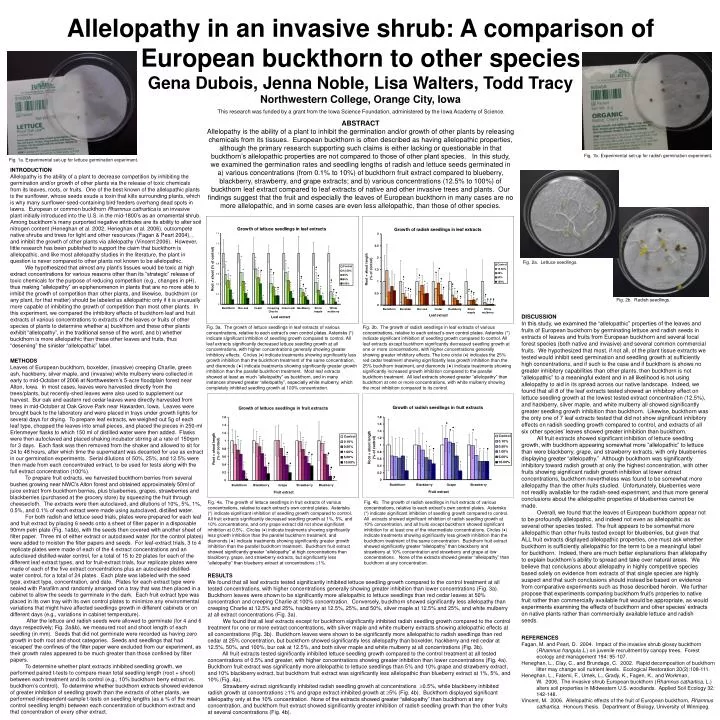
Allelopathy in an invasive shrub: A comparison of European buckthorn to other species Gena Dubois, Jenna Noble, Lisa Walters, Todd Tracy Northwestern College, Orange City, Iowa This research was funded by a grant from the Iowa Science Foundation, administered by the Iowa Academy of Science. ABSTRACT Allelopathy is the ability of a plant to inhibit the germination and/or growth of other plants by releasing chemicals from its tissues. European buckthorn is often described as having allelopathic properties, although the primary research supporting such claims is either lacking or questionable in that buckthorn’s allelopathic properties are not compared to those of other plant species. In this study, we examined the germination rates and seedling lengths of radish and lettuce seeds germinated in a) various concentrations (from 0.1% to 10%) of buckthorn fruit extract compared to blueberry, blackberry, strawberry, and grape extracts; and b) various concentrations (12.5% to 100%) of buckthorn leaf extract compared to leaf extracts of native and other invasive trees and plants. Our findings suggest that the fruit and especially the leaves of European buckthorn in many cases are no more allelopathic, and in some cases are even less allelopathic, than those of other species. Fig. 1b. Experimental set-up for radish germination experiment. Fig. 1a. Experimental set-up for lettuce germination experiment. INTRODUCTION Allelopathy is the ability of a plant to decrease competition by inhibiting the germination and/or growth of other plants via the release of toxic chemicals from its leaves, roots, or fruits. One of the best known of the allelopathic plants is the sunflower, whose seeds exude a toxin that kills surrounding plants, which is why many sunflower-seed-containing bird feeders overhang dead spots in lawns. European or common buckthorn Rhamnus cathartica is an invasive plant initially introduced into the U.S. in the mid-1800’s as an ornamental shrub. Among buckthorn’s many purported negative attributes are its ability to alter soil nitrogen content (Heneghan et al. 2002, Heneghan et al. 2006), outcompete native shrubs and trees for light and other resources (Fagan & Peart 2004), , and inhibit the growth of other plants via allelopathy (Vincent 2006). However, little research has been published to support the claim that buckthorn is allelopathic, and like most allelopathy studies in the literature, the plant in question is never compared to other plants not known to be allelopathic. We hypothesized that almost any plant’s tissues would be toxic at high extract concentrations for various reasons other than its “strategic” release of toxic chemicals for the purpose of reducing competition (e.g., changes in pH), thus making “allelopathy” an epiphenomenon in plants that are no more able to inhibit the growth of competition than other plants, and likewise, buckthorn (or any plant, for that matter) should be labeled as allelopathic only if it is unusually more capable of inhibiting the growth of competition than most other plants. In this experiment, we compared the inhibitory effects of buckthorn leaf and fruit extracts of various concentrations to extracts of the leaves or fruits of other species of plants to determine whether a) buckthorn and these other plants exhibit “allelopathy”, in the traditional sense of the word, and b) whether buckthorn is more allelopathic than these other leaves and fruits, thus “deserving” the sinister “allelopathic” label. Fig. 2a. Lettuce seedlings. Fig. 2b. Radish seedlings. DISCUSSION In this study, we examined the “allelopathic” properties of the leaves and fruits of European buckthorn by germinating lettuce and radish seeds in extracts of leaves and fruits from European buckthorn and several local forest species (both native and invasive) and several common commercial fruits. We hypothesized that most, if not all, of the plant tissue extracts we tested would inhibit seed germination and seedling growth at sufficiently high concentrations, and if such is the case and if buckthorn is shows no greater inhibitory capabilities than other plants, then buckthorn is not “allelopathic” to a meaningful extent and in all likelihood is not using allelopathy to aid in its spread across our native landscape. Indeed, we found that all 8 of the leaf extracts tested showed an inhibitory effect on lettuce seedling growth at the lowest tested extract concentration (12.5%), and hackberry, silver maple, and white mulberry all showed significantly greater seedling growth inhibition than buckthorn. Likewise, buckthorn was the only one of 7 leaf extracts tested that did not show significant inhibitory effects on radish seedling growth compared to control, and extracts of all six other species’ leaves showed greater inhibition than buckthorn. All fruit extracts showed significant inhibition of lettuce seedling growth, with buckthorn appearing somewhat more “allelopathic” to lettuce than were blackberry, grape, and strawberry extracts, with only blueberries displaying greater “allelopathy.” Although buckthorn was significantly inhibitory toward radish growth at only the highest concentration, with other fruits showing significant radish growth inhibition at lower extract concentrations, buckthorn nevertheless was found to be somewhat more allelopathy than the other fruits studied. Unfortunately, blueberries were not readily available for the radish-seed experiment, and thus more general conclusions about the allelopathic properties of blueberries cannot be made. Overall, we found that the leaves of European buckthorn appear not to be profoundly allelopathic, and indeed not even as allelopathic as several other species tested. The fruit appears to be somewhat more allelopathic than other fruits tested except for blueberries, but given that ALL fruit extracts displayed allelopathic properties, one must ask whether buckthorn is sufficiently allelopathic for the term to be a meaningful label for buckthorn. Indeed, there are much better explanations than allelopathy to explain buckthorn’s ability to spread and take over natural areas. We believe that conclusions about allelopathy in highly competitive species based solely on evidence from extracts of that single species are highly suspect and that such conclusions should instead be based on evidence from comparative experiments such as those described herein. We further propose that experiments comparing buckthorn fruit’s properies to native fruit rather than commercially available fruit would be appropriate, as would experiments examining the effects of buckthorn and other species’ extracts on native plants rather than commercially available lettuce and radish seeds. Fig. 3a. The growth of lettuce seedlings in leaf extracts of various concentrations, relative to each extract’s own control plates. Asterisks (*) indicate significant inhibition of seedling growth compared to control. All leaf extracts significantly decreased lettuce seedling growth at all concentrations, with higher concentrations generally showing greater inhibitory effects. Circles () indicate treatments showing significantly less growth inhibition than the buckthorn treatment of the same concentration, and diamonds () indicate treatments showing significantly greater growth inhibition than the parallel buckthorn treatment. Most leaf extracts showed at least as much “allelopathy” as buckthorn, and in many instances showed greater “allelopathy”, especially white mulberry, which completely inhibited seedling growth at 100% concentration. Fig. 3b. The growth of radish seedlings in leaf extracts of various concentrations, relative to each extract’s own control plates. Asterisks (*) indicate significant inhibition of seedling growth compared to control. All leaf extracts except buckthorn significantly decreased seedling growth at one or more concentrations, with higher concentrations generally showing greater inhibitory effects. The lone circle () indicates the 25% red cedar treatment showing significantly less growth inhibition than the 25% buckthorn treatment, and diamonds () indicate treatments showing significantly increased growth inhibition compared to the parallel buckthorn treatment. All leaf extracts showed greater “allelopathy” than buckthorn at one or more concentrations, with white mulberry showing the most inhibition compared to its control. METHODS Leaves of European buckthorn, boxelder, (invasive) creeping Charlie, green ash, hackberry, silver maple, and (invasive) white mulberry were collected in early to mid-October of 2006 at Northwestern’s 5-acre floodplain forest near Alton, Iowa. In most cases, leaves were harvested directly from the trees/plants, but recently-shed leaves were also used to supplement our harvest. Bur oak and eastern red cedar leaves were directly harvested from trees in mid-October at Oak Grove Park near Hawarden, Iowa. Leaves were brought back to the laboratory and were placed in trays under growth lights for several days for drying. To prepare leaf extracts, we weighed out 5g of each leaf type, chopped the leaves into small pieces, and placed the pieces in 250-ml Erlenmeyer flasks to which 150 ml of distilled water were then added. Flasks were then autoclaved and placed shaking incubator stirring at a rate of 150rpm for 3 days. Each flask was then removed from the shaker and allowed to sit for 24 to 48 hours, after which time the supernatant was decanted for use as extract in our germination experiments. Serial dilutions of 50%, 25%, and 12.5% were then made from each concentrated extract, to be used for tests along with the full extract concentration (100%). To prepare fruit extracts, we harvested buckthorn berries from several bushes growing near NWC’s Alton forest and obtained approximately 50ml of juice extract from buckthorn berries, plus blueberries, grapes, strawberries and blackberries (purchased at the grocery store) by squeezing the fruit through cheesecloth. The extracts were then autoclaved, and dilutions of 10%, 5%, 1%, 0.5%, and 0.1% of each extract were made using autoclaved, distilled water. For both radish and lettuce seed trials, plates were prepared for each leaf and fruit extract by placing 6 seeds onto a sheet of filter paper in a disposable 90mm petri plate (Fig. 1a&b), with the seeds then covered with another sheet of filter paper. Three ml of either extract or autoclaved water (for the control plates) were added to moisten the filter papers and seeds. For leaf-extract trials, 3 to 4 replicate plates were made of each of the 4 extract concentrations and an autoclaved distilled-water control, for a total of 15 to 20 plates for each of the different leaf extract types, and for fruit-extract trials, four replicate plates were made of each of the five extract concentrations plus an autoclaved distilled-water control, for a total of 24 plates. Each plate was labeled with the seed type, extract type, concentration, and date. Plates for each extract type were sealed with Parafilm and randomly arranged on a tray that was then placed in a cabinet to allow the seeds to germinate in the dark. Each fruit extract type was placed in its own tray with its own control plates to minimize any environmental variations that might have affected seedlings growth in different cabinets or on different days (e.g., variations in cabinet temperature). After the lettuce and radish seeds were allowed to germinate (for 4 and 6 days respectively; Fig. 2a&b), we measured root and shoot length of each seedling (in mm). Seeds that did not germinate were recorded as having zero growth in both root and shoot categories. Seeds and seedlings that had ‘escaped’ the confines of the filter paper were excluded from our experiment, as their growth rates appeared to be much greater than those confined by filter papers. To determine whether plant extracts inhibited seedling growth, we performed paired t-tests to compare mean total seedling length (root + shoot) between each treatment and its control (e.g., 10% buckthorn berry extract vs. buckthorn’s control). To determine whether buckthorn extracts showed evidence of greater inhibition of seedling growth than the extracts of other plants, we performed independent-sample t-tests on seedling lengths (as a % of the mean control seedling length) between each concentration of buckthorn extract and that concentration of every other extract. Fig. 4a. The growth of lettuce seedlings in fruit extracts of various concentrations, relative to each extract’s own control plates. Asterisks (*) indicate significant inhibition of seedling growth compared to control. All fruit extracts significantly decreased seedling growth at 1%, 5%, and 10% concentrations, and only grape extract did not show significant inhibition at 0.5%. Circles () indicate treatments showing significantly less growth inhibition than the parallel buckthorn treatment, and diamonds () indicate treatments showing significantly greater growth inhibition than the parallel buckthorn treatment. Buckthorn fruit extract showed significantly greater “allelopathy” at high concentrations than blackberry, grape, and strawberry extracts, but significantly less “allelopathy” than blueberry extract at concentrations 1%. Fig. 4b. The growth of radish seedlings in fruit extracts of various concentrations, relative to each extract’s own control plates. Asterisks (*) indicate significant inhibition of seedling growth compared to control. All extracts showed significant inhibition of radish seedling growth at 10% concentration, and all fruits except buckthorn showed significant inhibition for at least one of the intermediate concentrations. Circles () indicate treatments showing significantly less growth inhibition than the buckthorn treatment of the same concentration. Buckthorn fruit extract showed significantly greater “allelopathy” than blackberry and strawberry at 10% concentration and strawberry and grape at low concentrations. None of the extracts showed greater “allelopathy” than buckthorn at any concentration. RESULTS We found that all leaf extracts tested significantly inhibited lettuce seedling growth compared to the control treatment at all tested concentrations, with higher concentrations generally showing greater inhibition than lower concentrations (Fig. 3a). Buckthorn leaves were shown to be significantly more allelopathic to lettuce seedlings than red cedar leaves at 50% concentration and creeping Charlie at 100% concentration. Conversely, buckthorn showed significantly less allelopathy than creeping Charlie at 12.5% and 25%, hackberry at 12.5%, 25%, and 50%, silver maple at 12.5% and 25%, and white mulberry at all extract concentrations (Fig. 3a). We found that all leaf extracts except for buckthorn significantly inhibited radish seedling growth compared to the control treatment for one or more extract concentrations, with silver maple and white mulberry extracts showing allelopathic effects at all concentrations (Fig. 3b). Buckthorn leaves were shown to be significantly more allelopathic to radish seedlings than red cedar at 25% concentration, but buckthorn showed significantly less allelopathy than boxelder, hackberry and red cedar at 12.5%, 50%, and 100%, bur oak at 12.5%, and both silver maple and white mulberry at all concentrations (Fig. 3b). All fruit extracts tested significantly inhibited lettuce seedling growth compared to the control treatment at all tested concentrations of 0.5% and greater, with higher concentrations showing greater inhibition than lower concentrations (Fig. 4a). Buckthorn fruit extract was significantly more allelopathic to lettuce seedlings than 5% and 10% grape and strawberry extract, and 10% blackberry extract, but buckthorn fruit extract was significantly less allelopathic than blueberry extract at 1%, 5%, and 10% (Fig. 4a). Strawberry extract significantly inhibited radish seedling growth at concentrations 0.5%, while blackberry inhibited radish growth at concentrations 1% and grape extract inhibited growth at 5% (Fig. 4b). Buckthorn displayed significant allelopathy only at the 10% concentration. None of the extracts showed greater “allelopathy” than buckthorn at any concentration, and buckthorn fruit extract showed significantly greater inhibition of radish seedling growth than the other fruits at several concentrations (Fig. 4b). REFERENCES Fagan, M. and Peart, D. 2004. Impact of the invasive shrub glossy buckthorn (Rhamnus frangula L.) on juvenile recruitment by canopy trees. Forest ecology and management 194: 95-107. Heneghan, L., Clay, C., and Brundage, C. 2002. Rapid decomposition of buckthorn litter may change soil nutrient levels. Ecological Restoration 20(2):108-111. Heneghan, L., Fatemi, F., Umek, L., Grady, K., Fagen, K., and Workman, W. 2006. The invasive shrub European buckthorn (Rhamnus cathartica, L.) alters soil properties in Midwestern U.S. woodlands. Applied Soil Ecology 32: 142-148. Vincent, M. 2006. Allelopathic effects of the fruit of European buckthorn, Rhamnus cathartica. Honours thesis. Department of Biology, University of Winnipeg.