Download
1 / 20
240 likes | 506 Views
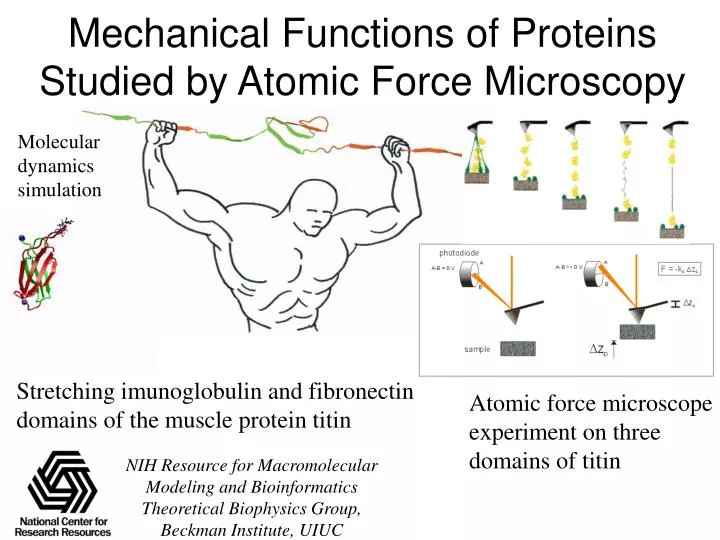
Mechanical Functions of Proteins Studied by Atomic Force Microscopy. Molecular dynamics simulation. C. D. F. E. G. Stretching imunoglobulin and fibronectin domains of the muscle protein titin. Atomic force microscope experiment on three domains of titin. B. A.

E N D
Mechanical Functions of Proteins Studied by Atomic Force Microscopy Molecular dynamics simulation C D F E G Stretching imunoglobulin and fibronectin domains of the muscle protein titin Atomic force microscope experiment on three domains of titin B A NIH Resource for Macromolecular Modeling and Bioinformatics Theoretical Biophysics Group, Beckman Institute, UIUC
Forces can be substrates, products, signals, catalysts of cellular processes But to what degree can proteins and DNA sustain forces? How do proteins need to be designed to build machines from them?
Titin the Longest Protein in the Human Genome I1 I27 titin I-band sarcomere 2 m I1 I41 PEVK region; random coil I27 Under weak forces, the PEVK region extends (entropic spring) titin immunoglobulin-like (Ig) domain Under intermediate forces, Ig domains stretch by ~10A; under strong forces, they unfold one by one, by 250A 40 Å
Atomic Force Microscope Instrument 15 cm
v AFM Studies of Titin And FN-III Modules AFM tip • Forced unfolding of identical modules occurs one by one Titin I27 FN-III 600 Force (pN) Force (pN) 400 0 0 50 100 150 Extension (nm) Extension (nm) Reviewed in Fisheret al. Nature Struct. Biol.7:719-724 (2001)
Stretching modular proteins – Detailed View Extension occurs in two steps extension Schematic view and typical Extension vs. force plot Distribution of measured forces For step 1 and step 2
Probing the Passive Elasticity of Muscle: I27 Marszalek et al., Nature, 402,100-103 (1999) Krammer et al., PNAS, 96, 1351-1356 (1999) Lu et al., Biophys. J., 75, 662-671 (1998) sarcomere Computational Stretching of Ig: Trajectory Animation Gaub et al., Fernandez et al. Hui Lu, Barry Isralewitz titin immunoglobulin-like (Ig) domain
Two Force Bearing Components Two types of SMD simulations I. Native structure (2 A of extension) II. Mechanical unfolding intermediate (10 A of extension) III. Unfolded state (250 A of extension) Constant velocity: Lu et al, Biophys J., 72, 1568 (1007) six interstrand (A’ – G) hydrogen bonds three interstrand (A – B) hydrogen bonds force Constant force: Lu and Schulten, Chem. Phys., 247 (1999) 141. Reviewed inIsralewitzet al. Curr. Opinion Struct. Biol. 11:224-230 (2001) Hui Lu, Barry Isralewitz
Quantitative Comparison AFM data SMD data Extrapolation of AFM data Bridging the gap between SMD and AFM experiments Steered Molecular Dynamics (SMD) Force-extension curve Force-pulling velocity relationship AFM range Current SMD range Target simulation range Extension curve Hui Lu, Barry Isralewitz
Titin Ig Mechanical Unfolding Intermediate SMD simulations with constant forces AFM force-extension profile E6P Marszalek, Lu, Li, Carrion, Oberhauser, Schulten, and Fernandez, Nature, 402, 100 (1999). Hui Lu
Sampling Titin Ig Unfolding Barrier U(x) DU -Fx (F fixed) a a b b Lu and Schulten, Chem. Phys., 247 (1999) 141. The key event of titin Ig unfolding can be examined as a barrier crossing process. t(F) = 2tDd(F) [ed(F) – d(F) –1] t(D) = (b – a)2/2D ~ 25 ns d(F) = b [DU – F(b-a)] determination of barrier height based on mean first passage time 1200 the best fit suggests a potential barrier of D 800 U = 20 kcal/mol burst time (ps) 400 0 400 600 800 1000 1200 0 stationary force applied (pN) Chemical denaturing experiments (Carrion-Vazquez et al., PNAS, 1999) show a similar U = 22 kcal/mol. Hui Lu NIH Resource for Macromolecular Modeling and Bioinformatics Theoretical Biophysics Group, Beckman Institute, UIUC
Sampling Distribution of Barrier Crossing Times Fraction N(t) of domains that have not crossed the barrier, for linear model potential, is: Approximated by double exponential (holds for arbitrary potential): N(t) = [t1 exp(-t/t1) – t2 exp(-t/t2)]/(t1-t2), Nadler & Schulten, JCP., 82, 151-160 (1985) Multiple runs of I27 unfolding with a 750 pN stationary force Barrier crossing times of 18 SMD simulations Theoretical prediction of the barrier crossing times U = 20 kcal/mol Chemical denaturing experiments (Carrion-Vazquez et al., PNAS, 1999) show a similar U = 22 kcal/mol. Hui Lu NIH Resource for Macromolecular Modeling and Bioinformatics Theoretical Biophysics Group, Beckman Institute, UIUC
Rupture/Unfolding Force F0 and its Distribution 1200 the best fit suggests a potential barrier of t(F0) = 1 ms time of measurement => F0 rupture/unfolding force D 800 U = 20 kcal/mol burst time (ps) 400 Distribution of rupture/unfolding force 0 400 600 800 1000 1200 stationary force applied (pN) k = d2(F)/2tDkv Israilev et al., Biophys. J., 72, 1568-1581 (1997) Balsera et al., Biophys. J., 73, 1281-1287 (1997) Fernandez et al., (PNAS, in press) Manel Balsera, Sergei Izrailev NIH Resource for Macromolecular Modeling and Bioinformatics Theoretical Biophysics Group, Beckman Institute, UIUC
Water-Backbone Interactions Control Unfolding 1000 pN 750 pN B A G A' water greases protein folding ! During stretching, water molecules attack repeatedly interstrand hydrogen bonds, about 100 times / ns; for forces of 750 pN water fluctuation controls unfolding! E12 Lu and Schulten, Biophys J.79: 51-65 (2000)
Titin Domain Heterogenity: pre-stretching slack tight in I1 disulfide bond Hydrogen bond energies /(Kcal/mol) I1 in 70 Å water sphere of 70 Å (18,000 atoms) K6(H)-V29(O) E3 K31 E3 K31 E3 K31 A Use of program NAMD on 32 Processor (1.1 GHz Athlon) Linux cluster: 1 day/ns A B A B A B E3(H)-K31(O) E3(O)-K31(H) B V29 K6 V29 K6 V29 K6 K6(O)-V29(H) F8(H)-R27(O) E9(O)-R27(H) F8 G F8 R27 R27 F8 A’ R27 E9 E9 E9 F time / ps I1 I27 PEVK 3 A-B bonds 6 A-B bonds SMD-CF-750 pN easy slack tight slack 4 H bonds Hydrogen Bond Energy (kcal/mol) Extension (Å) 2 H bonds titin I1 titin I27 Mayanset al. Structure9:331-340 (2001) Time (ps) water native 410 ps, 8 Å 585 ps, 12 Å Mu Gao
B F A G Architecture and Function of Fibronectin Modules Extracellular matrix Fibronectin FN-I FN-II FN-III 10 m FN-III9 FN-III7 FN-III8 FN-III10 • Fibronectin is a major component of extracellular matrix • It consists of three types of modules: type I, type II, and type III • Highly extensible 35 Å Andre Krammer, U. Wash.
Fibronectin Matrix in Living Cell Culture FN-III Force (pN) Extension (nm) Atomic force microscopy observations 100 m Ohashi et al. Proc. Natl. Acad. Sci USA96:2153-2158 (1999)
Krammeret al. Proc. Natl. Acad. Sci USA 96:1351-1356 (1999) RGD Loop of FnIII10 Gly79 Asp80 Arg78 Val1C Thr94C Native FnIII10 fibronectin binding with RGD loop to integrin F= 500 pN Extension of 13 Å integrin interacting with extracellular matrix Andre Krammer, U. Washington http://www.ks.uiuc.edu
F Native Fibronectin III10 Probing Unfolding Intermediates in FN-III10 FnIII10 module solvated in a water box 5560367 Å3 (126,000 atoms) Steered Molecular Dynamics, periodic boundary conditions, NpT ensemble, Particle Mesh Edwald for full electrostatics NAMD on Linux cluster of 32 Athlon 1.1GHz processors: 10 days/ns Mu Gao
ATOMIC FORCE MICROSCOPY FOR BIOLOGISTS by V J Morris, A R Kirby & A P Gunning 352pp Pub. date: Dec 1999 ISBN 1-86094-199-0 US$51 / £32 Contents: Apparatus Basic Principles Macromolecules Interfacial Systems Ordered Macromolecules Cells, Tissue and Biominerals Other Probe Microscopes Atomic force microscopy (AFM) is part of a range of emerging microscopic methods for biologists which offer the magnification range of both the light and electron microscope, but allow imaging under the 'natural' conditions usually associated with the light microscope. To biologists AFM offers the prospect of high resolution images of biological material, images of molecules and their interactions even under physiological conditions, and the study of molecular processes in living systems. This book provides a realistic appreciation of the advantages and limitations of the technique and the present and future potential for improving the understanding of biological systems.





















