Download

1 / 43
430 likes | 661 Views
Cytokines Online Pathfinder Encyclopaedia. http://www.copewithcytokines.de/cope.cgi

E N D
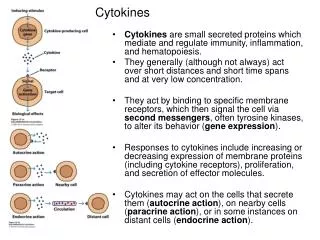
Cytokines Online Pathfinder Encyclopaedia • http://www.copewithcytokines.de/cope.cgi • The term cytokine, or Immunocytokines, was used initially to separate a group of immunomodulatory proteins, called also Immunotransmitters , from other Growth factors that modulate the proliferation and bioactivities of non-immune cells. • Some cytokines are produced by a rather limited number of different cell types while others are produced by almost the entire spectrum of known cell types. For lecture only; BC Yang
Cyto: cell -kines: kinesis, 可移動, 分裂 • In the more restricted sense cytokines comprise Interleukins , initially thought to be produced exclusively by leukocytes, Lymphokines , initially thought to be produced exclusively by lymphocytes, Monokines , initially thought to be produced exclusively by monocytes, interferons, initially thought to be involved in antiviral responses, colony stimulating factors, initially thought to support the growth of cells in semi-solid media, Chemokines , thought to be involved in Chemotaxis , and a variety of other proteins. For lecture only; BC Yang
Established cell lines that entirely depend for their survival and proliferation on the continuous presence of one or more of growth factors . Factor-dependent cell lines are capable of continuous growth in the presence of the growth factors and cytokines they require For lecture only; BC Yang
Hematopoiesis: Greek haima for blood and poiein, to make • Hematopoiesis is the dynamic and complex developmental process of the formation of new blood cells. • An early intra-embryonic site of hematopoiesis is found in the Paraaortic splanchnopleura and in a structure termed AGM ( abbr. for aorta, gonads, and mesonephros) ( Nishikawa et al). mice and birds • Hematopoiesis taking place prior to the development of the fetal liver is referred to as primitive hematopoiesis . The fetal liver is the site of definitive hematopoiesis early during embryonal development. • The bone marrow with its intersinusoidal spaces is the site responsible for the generation of blood cells in the post-natal phase. For lecture only; BC Yang
Hematopoesis www.wsu.edu/~ms523/ ms523s198.html • Sites of blood cell formation • Embryo: yolk sac • Fetus: spleen and liver • Adult: “bone marrow” sternum, ribs, pelvis, cranium, vertebrae, long bones (tibia and femur) For lecture only; BC Yang
The high proliferative potential colony assay • Bone marrow cells 25000 cells of Balb/c, after 150 mg/kg of 5-Fu. • Culture s are incubated under low oxygen tension for 12 days and scored for HPP-CFU. For lecture only; BC Yang
Moore, 10:2719 The development of neutrophils from the stem cell stage For lecture only; BC Yang
http://www.whfreeman.com/immunology/CH03/hematopoiesis.htm For lecture only; BC Yang
http://www.whfreeman.com/immunology/CH03/hematopoiesis.htm In the presence of infection, cytokines produced by activated macrophages and T-helper cells induce additional hematopoietic activity, resulting in rapid expansion of white blood cells that participate in fighting infection. For lecture only; BC Yang
Mosmann TR et al , J Immunol (1986) 136:2348-2357 • This is underscored by the uniform phenotype of our anti-CRBC T cell lines, which were all Th1. Only when we examined T cell clones specific for FGG or alloantigens did we discover antigen-specific T cells of the Th2 cells • Th2 cells produce activities that strongly enhance IgE/IgG1 production....., whereas Th1 cells produce interferon-g, which strongly inhibits the enhancing activities in Th2 supernatants ..... For lecture only; BC Yang
Two types of murine helper T cell clone Mosmann TR et al , J Immunol (1986) 136:2348-2357 For lecture only; BC Yang
Cytokine secretion phenotypes of mouse T cells For lecture only; BC Yang
Immunosuppresive cytokines For lecture only; BC Yang
Th1 and Th2 cell generation is regulated by the equilibrium between different groups of cytokines, with dominant effects of IL-12 and IL-4 (with feed back mechanisms) For lecture only; BC Yang
The commitment of TH0 cells to become TH1 or TH2 is influenced by cytokines secreted by the 2 subtypes themselves and by macrophages , NK cells and mast cells. http://www.brown.edu/Courses/Bio_160/Projects1999/ies/cytok.html For lecture only; BC Yang
Cytokine soup during disease For lecture only; BC Yang
A Th1 to Th2 switch ia a critical step in the etiology of HIV infectionClerici M and Shearer GM, Immunol Today 14:107-111 (1993) For lecture only; BC Yang
Becoming a paradigm for disease management Susceptibility to Leishmania major infection in interleukin-4-deficient mice. Noben-Trauth et al 1996, Science 271:987-989 For lecture only; BC Yang
Induction of Th1 and Th2 responses: a key role for the natural immune respones? Immunol Today 13:379-381 (1992) For lecture only; BC Yang
Role of IL-12 in the generation of Th1 cells For lecture only; BC Yang
Cytokine responsiveness in a human volunteer injected with LPS. TNF-a rises almost immediately and peaks at 1.5 hours; the sharp decline of TNF-a may be due to modulation by its soluble receptor sTNF-R. A second wave of cytokines that peaks at 3 hours activates the acute phase response in the liver and the systemic pituitary response (both via IL-6) and the activation and chemotaxis of neutrophils (via IL-6, IL-8 and G-CSF). Pituitary-derived adrenocorticotropic hormone (ACTH) and migration inhibition factor (MIF) peak at 5 hours and coincide with peak levels of the regulatory cytokines IL-Ra and IL-10 that counteract the release or activity of proinflammatory cytokines. For lecture only; BC Yang
IL-4 instructs TH1 responses and resistance to Leishmania major in susceptible BALB/c miceBiedermann T et al. 2001, Nature Immunol 2:11 As usual, things may not be so simple • When present during the initial activation of dendritic cells (DCs) by infectious agents, IL-4 instructed DCs to produce IL-12 and promote TH1 development. This TH1 response established resistance to Leishmania major in susceptible BALB/c mice. When present later, during the period of T cell priming, IL-4 induced TH2 differentiation and progressive leishmaniasis in resistant mice. Because immune responses developed via the consecutive activation of DCs and then T cells, the contrasting effects of IL-4 on DC development and T cell differentiation led to immune responses that had opposing functional phenotypes. For lecture only; BC Yang
IL-4 instructs DCs to produce increased amounts of IL-12 and to promote TH1 development in vitro. For lecture only; BC Yang
IL-4 instructs resistance to L. major in susceptible BALB/c mice. Groups of five BALB/c mice or more were inoculated in one hind footpad with 2×105 stationary phase L. major promastigotes either of strain LV39, MRHO/Sv/59/P (a) or strain MHOM/IL/81/FE/BNI (b). For lecture only; BC Yang
IL-4 instructs parasite containment in susceptible BALB/c mice.BALB/c mice were infected with L. major and treated twice with the indicated doses of IL-4 or NMS during the first 8 h of infection. After 9 weeks, the number of viable parasites in infected tissues was estimated using a parasite-limiting dilution assay. For lecture only; BC Yang
IL-4 treatment during the first 8 h of L. major infection instructs IL-12–producing DC1s and suppresses IL-4 expression. Control mice (Control and IL-4) or mice infected with L. major promastigotes (3×106) were treated either with NMS or twice with 1.0 μg of IL-4 at 0 and 8 h of infection. For lecture only; BC Yang
Induction of TH1 responses with IL-4 depends on IL-12. BALB/c mice were infected with L. major and treated twice with IL-4 (at 0 and 8 h) and either NMS or anti–IL-12. After 8 weeks, popliteal lymph node cells were isolated, RNA extracted and expression of IL-4 and IFN-γ mRNA determined by semi-quantitative RT-PCR. (a) The relative changes in IL-4 and IFN-γ mRNA were normalized to the values in control infected BALB/c mice, arbitrarily fixed as 100. (b) The ratio of IL-4:IFN-γ mRNA for each experimental group was determined. L. major infection, cytokine treatment and mRNA analysis were done. For lecture only; BC Yang
Treatment with anti–IL-12 abrogates IL-4–induced resistance to L. major in BALB/c mice. Mice were infected with L. major promastigotes and treated with either NMS (open circles) or twice with 1.0 μg of IL-4 (at 0 and 8 h) and either NMS (filled squares) or the IL-12 mAbs C17.8 and C17.15 (open triangles) 12 and 3 h before infection. One of two similar experiments is shown. Infection, cytokine treatment and weekly monitoring of footpad lesions were done . For lecture only; BC Yang
Extension of IL-4 treatment into the period of T cell priming reverses IL-4–induced resistance to L. major. Three groups of mice were infected and treated with either 1.0 μg of IL-4 or NMS. The first L. major–infected group received NMS (open circles), the second 1.0 μg of IL-4 at 0 and 8 h (filled squares) and the third 1.0 μg of IL-4 at 0, 8, 16 and 24 h (crosses). Infection, cytokine treatment and weekly monitoring of footpad lesions were done . For lecture only; BC Yang
Timing determines whether IL-4 treatment instructs a TH1 phenotype and resistance or a TH2 phenotype and susceptibility to L. major in TCR Vβ4-deficient mice. (a,b) After parasite inoculation,Vβ4-deficient BALB/c mice were treated during the first 64 h with either eight injections of PBS–1% NMS or eight injections of 0.1 μg of IL-4 (total amount: 0.8 μg). Alternatively, mice were treated twice during the first 8 h of infection with 1.0 μg of IL-4 as in Fig. 2. Eight weeks after infection, cells from draining lymph nodes were stimulated in vitro with ultraviolet-irradiated parasites and cytokines were determined in the supernatants as described (see Methods). The IL-4:IFN-γ ratio was 3.71 in mice that received IL-4 over 64 h and 0.37 in mice that received two injections of IL-4 at 0 h and 8 h. (c). For lecture only; BC Yang
How Th1/2 cytokines are regulated? • Transcriptional regulation of Th1/Th2 polarization, Jyothi Rengarajan J et al 2000, Immunol Today 21:479 The two polarized T helper (Th) subsets Th1 and Th2 are identified by their signature cytokines, interferon-g (IFN-g) and interleukin 4 (IL-4) respectively. Understanding the transcriptional regulation of cytokine expression is therefore critical for elucidating the process of Th cell differentiation. Ubiquitous and tissue-specific transcription factors, as well as chromatin remodeling of genomic loci have been implicated in IL-4 and IFN-g regulation. For lecture only; BC Yang
Class I cytokine receptor family (Hematopoietin recetor) • this family includes receptors for growth hormone and prolactin. There are conserved amino acid sequence motifs in the extracellular domain - 4 positionally conserved cysteine residues (CCCC) and a conserved sequence of Trp-Ser-X-Trp-Ser (WSXWS) where X is a nonconserved amino acid. The receptors consist of 2 polypeptide chains: a cytokine-specific subunit and a signal-transducing subunit which is usually not specific for the cytokine. http://www.umdnj.edu/pathnweb/genpath/lec_1/Class_I_Cytokine_Receptors/class_i_cytokine_receptors.htm For lecture only; BC Yang
Class II cytokine receptor family(Interferon receptor family) • The ligands for these receptors are the three interferons a, ß, and g. These receptors possess the conserved cysteine motifs, but lack the WSXWS motif present in class I cytokine receptors. For lecture only; BC Yang
Overview of Th cell differentiation. A naive CD4 T cell is activated via the TCR when it encounters antigen presented by an antigen presenting cell. For lecture only; BC Yang
Regulation at transcriptional level • GATA3, a zinc finger protein that was originally identified as binding the TCRa gene enhancer via a WGATAR sequence. • T-bet (T-box expressed in T cells) is a member of the T-box family of transcription factors that regulate several developmental processes. T-bet expression strongly correlates with IFN-g expression. For lecture only; BC Yang
(a) IL-4 regulatory regions: the IL-4 proximal promoter with cis and trans elements and map of IL-4 locus showing DNAse I HSs. • (b) IFN-g regulatory region and map of locus with DNAse I HS sites. For lecture only; BC Yang
Striking a balance: GATA3 and T-bet – model for Th1/Th2 polarization. Signals through the TCR and cytokine receptors can lead to the initiation of a Th1 program (via Stat4 activation) and the induction of T-bet, which promotes Th1 lineage commitment. Signals that favor the activation of Stat6 induce GATA3 leading to Th2 differentiation. c-Maf is then upregulated leading to increased IL-4 production and Th2 polarization. For lecture only; BC Yang
More than immunoregulatroy actions • Cytokine regulation of neuronal differentiation of hippocampal progenitor cells (Mehler MF et al. 1993, Nature 362:62-65.) • The signalling mechanisms governing haematolymphopoiesis and those regulating neural development may be closely related, as indicated by similarities of higher-order structure and function of the cytokines involved, of the regional and temporal regulation of their transcription and translation, and of their bioactivity. • The mechanisms regulating lineage commitment and cellular differentiation in the neural and haematopoietic systems are similar. For lecture only; BC Yang
Electrophysiological analysis of the effects of growth factor treatment on neuronal maturation For lecture only; BC Yang
Morphological and immunocytochemical indices of progressive neuronal maturation For lecture only; BC Yang
Immune System and the BrainEstrogen, microglia and the IBB The microglial-cytokine--estrogen mediated network is described as a major player in the brain including the Immune Brain Barrier. One of the functional areas involved maybe the role that microglia could play in synaptic plasticity. http://info.med.yale.edu/obgyn/reproimmuno/projects/immune.html For lecture only; BC Yang
Combinatorial signal by inflammatory cytokines and chemokines mediate leukocyte interactions with extracellular matrix Vaday GG et al. 2001, J leukoc Biol 69:885-892 For lecture only; BC Yang
limitations of cytokines for therapeutic use • High local concentrations of cytokines during immune response cannot be mimicked clinically • Short half-life of cytokines • Pleiotropic effects can cause unpredictable side-effects For lecture only; BC Yang