Download
1 / 1
10 likes | 98 Views
Chimeric HIV-1 viruses as a research tool. Leo Heyndrickx, Ellen Van Gulck , Lotte Bracke, Kevin Ariën , Rafael Van den Bergh and Guido Vanham Virology unit, Department of Microbiology, Institute of Tropical Medicine (ITM), Antwerp, Belgium. . Knowledge for Growth , Gent, 05.05.2011.

E N D
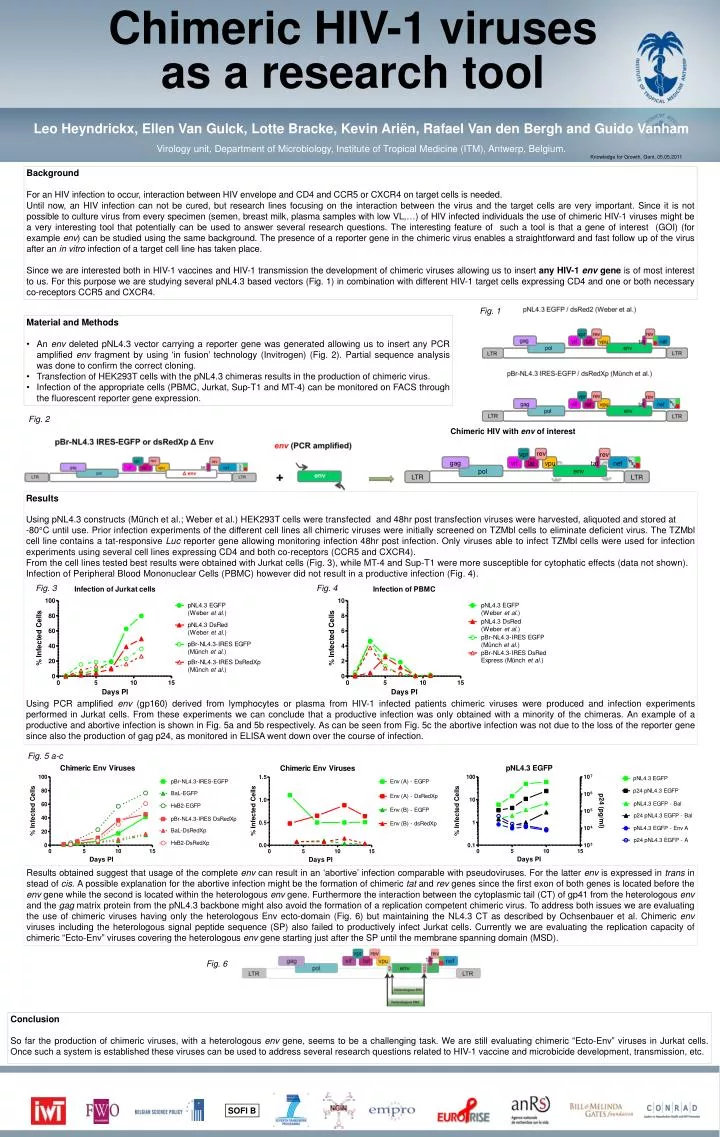
Chimeric HIV-1 viruses as a research tool Leo Heyndrickx, Ellen Van Gulck, Lotte Bracke, Kevin Ariën, Rafael Van den Bergh and Guido Vanham Virology unit, Department of Microbiology, Institute of Tropical Medicine (ITM), Antwerp, Belgium. Knowledge forGrowth, Gent, 05.05.2011 Background For an HIV infection to occur, interaction between HIV envelope and CD4 and CCR5 or CXCR4 on target cells is needed. Until now, an HIV infection can not be cured, but research lines focusing on the interaction between the virus and the target cells are very important. Since it is not possible to culture virus from every specimen (semen, breast milk, plasma samples with low VL,…) of HIV infected individuals the use of chimeric HIV-1 viruses might be a very interesting tool that potentially can be used to answer several research questions. The interesting feature of such a tool is that a gene of interest (GOI) (for example env) can be studied using the same background. The presence of a reporter gene in the chimeric virus enables a straightforward and fast follow up of the virus after an in vitro infection of a target cell line has taken place. Since we are interested both in HIV-1 vaccines and HIV-1 transmission the development of chimeric viruses allowing us to insert any HIV-1 env gene is of most interest to us. For this purpose we are studying several pNL4.3 based vectors (Fig. 1) in combination with different HIV-1 target cells expressing CD4 and one or both necessary co-receptors CCR5 and CXCR4. Fig. 1 • Material and Methods • An env deleted pNL4.3 vector carrying a reporter gene was generated allowing us to insert any PCR amplified env fragment by using ‘in fusion’ technology (Invitrogen) (Fig. 2). Partial sequence analysis was done to confirm the correct cloning. • Transfection of HEK293T cells with the pNL4.3 chimeras results in the production of chimeric virus. • Infection of the appropriate cells (PBMC, Jurkat, Sup-T1 and MT-4) can be monitored on FACS through the fluorescent reporter gene expression. Fig. 2 Results Using pNL4.3 constructs (Münch et al.; Weber et al.) HEK293T cells were transfected and 48hr post transfection viruses were harvested, aliquoted and stored at -80°C until use. Prior infection experiments of the different cell lines all chimeric viruses were initially screened on TZMbl cells to eliminate deficient virus. The TZMbl cell line contains a tat-responsive Luc reporter gene allowing monitoring infection 48hr post infection. Only viruses able to infect TZMbl cells were used for infection experiments using several cell lines expressing CD4 and both co-receptors (CCR5 and CXCR4). From the cell lines tested best results were obtained with Jurkat cells (Fig. 3), while MT-4 and Sup-T1 were more susceptible for cytophatic effects (data not shown). Infection of Peripheral Blood Mononuclear Cells (PBMC) however did not result in a productive infection (Fig. 4). Using PCR amplified env (gp160) derived from lymphocytes or plasma from HIV-1 infected patients chimeric viruses were produced and infection experiments performed in Jurkat cells. From these experiments we can conclude that a productive infection was only obtained with a minority of the chimeras. An example of a productive and abortive infection is shown in Fig. 5a and 5b respectively. As can be seen from Fig. 5c the abortive infection was not due to the loss of the reporter gene since also the production of gag p24, as monitored in ELISA went down over the course of infection. Transmission Fig. 3 Fig. 4 Fig. 5 a-c Results obtained suggest that usage of the complete env can result in an ‘abortive’ infection comparable with pseudoviruses. For the latter env is expressed in trans in stead of cis. A possible explanation for the abortive infection might be the formation of chimeric tat and rev genes since the first exon of both genes is located before the env gene while the second is located within the heterologous env gene. Furthermore the interaction between the cytoplasmic tail (CT) of gp41 from the heterologous env and the gag matrix protein from the pNL4.3 backbone might also avoid the formation of a replication competent chimeric virus. To address both issues we are evaluating the use of chimeric viruses having only the heterologous Envecto-domain (Fig. 6) but maintaining the NL4.3 CT as described by Ochsenbauer et al. Chimeric env viruses including the heterologous signal peptide sequence (SP) also failed to productively infect Jurkat cells. Currently we are evaluating the replication capacity of chimeric “Ecto-Env” viruses covering the heterologous env gene starting just after the SP until the membrane spanning domain (MSD). Fig. 6 Conclusion So far the production of chimeric viruses, with a heterologous env gene, seems to be a challenging task. We are still evaluating chimeric “Ecto-Env” viruses in Jurkat cells. Once such a system is established these viruses can be used to address several research questions related to HIV-1 vaccine and microbicide development, transmission, etc. SOFI B






















