Download

1 / 50
500 likes | 520 Views
Learn about the evolution of metabolic pathways, enzyme organization, thermodynamics, ATP chemistry, and the interconversions among nucleotide phosphates in Glycolysis. This comprehensive guide dives into the practical aspects of biochemical thermodynamics and the compartmentation of pathways in different tissues.

E N D
Metabolism II:Glycolysis Andy HowardBiochemistry Lectures, Fall 201017 October 2010 Metabolism; Glycolysis
Metabolism Evolution of pathways Compartmentation Thermodynamics ATP High-energy compounds Redox and Energy Glycolysis Overview Steps through TIM Steps to pyruvate What we’ll discuss Metabolism; Glycolysis
Logistics • Midterm answer key will be posted as soon as I’m sure that everyone has taken the exam • Grading should be finished by Saturday • Next Monday’s class (10/25) will be held on Thursday morning; details to follow Metabolism; Glycolysis
Evolution of pathways:how have new pathways evolved? • Add a step to an existing pathway • Evolve a branch on an existing pathway • Backward evolution • Duplication of existing pathway to create related reactions • Reversing an entire pathway Metabolism; Glycolysis
Adding a step Original pathway E1 E2 E3 E4 E5 A B C D E P • When the organism makes lots of E, there’s good reason to evolve an enzyme E5 to make P from E. • This is how asn and gln pathways (from asp & glu) work Metabolism; Glycolysis
Evolving a branch E3 E1 E2 • Original pathway: D A B C X • Fully evolved pathway: D A B C X E3a E3b Metabolism; Glycolysis
Backward evolution • Original system has lots of E P • E gets depleted over time; • need to make it from D, • so we evolve enzyme E4 to do that. • Then D gets depleted; • need to make it from C, • so we evolve E3 to do that • And so on Metabolism; Glycolysis
Duplicated pathways • Homologous enzymes catalyze related reactions;this is how trp and his biosynthesis enzymes seem to have evolved • Variant: recruit some enzymes from another pathway without duplicating the whole thing (example: ubiquitination) Metabolism; Glycolysis
Reversing a pathway • We’d like to think that lots of pathways are fully reversible • Usually at least one step in any pathway is irreversible (Go’ < -15 kJ mol-1) • Say CD is irreversible so E3 only works in the forward direction • Then D + ATP C + ADP + Pi allows us to reverse that one step with help • The other steps can be in common • This is how glycolysis evolved from gluconeogenesis Metabolism; Glycolysis
Enzyme organization • Enzymatic reactions are organized into pathways, where reactions proceed in an ordered sequence leading from the first reactant to the final product • Often, especially in eukaryotes, the relevant enzymes are spatially organized into groupings that allow one enzyme to emit its product in a position where it can be immediately picked up as a substrate by the next enzyme (G&G fig. 17.5) • These grouped enzymes are often membrane-bound to provide physical stability • Metabolons are stable multienzyme complexes that work this way Metabolism; Glycolysis
Compartmentation I:Localized pathways • Some are in membranes, some free in cytosol or in aqueous organelles • Several catabolic pathways in eukaryotes are localized in mitochondria • Corresponding anabolic pathways are in cytosol • Reduces likelihood of futile cycling • Multienzyme complexes, especially in eukaryotes • Provide entropic advantage • Often membrane-associated Metabolism; Glycolysis
Compartmentation II:Tissue Specialization • Obvious in multicellular eukaryotes • Individual cells may perform a limited number of metabolic roles • Some fully mature cells are anuclear • Requires careful cell-cell communication • Even in cyanobacteria Metabolism; Glycolysis
Thermodynamics • We did this carefully earlier:this is just a reminder • Remember that G is not Go’:G = Go’ + RT ln[products]/[reactants] • At equilibrium G = 0,so we can use that equation to find Go’ Metabolism; Glycolysis
Practical biochemical thermodynamics • Most reactions are considered either irreversible or fully reversible • Irreversible means Go’ < -20 kJ/mol; • Even with substrate and product concentrations considered, the reaction proceeds in only one direction • Reversible: -15 < Go’ < 15 butG very close to zero Metabolism; Glycolysis
ATP • Both the anhydride bonds are considered high-energy bonds; the phosphoester bond is not. • Remember the story about pyrophosphate!:PPi + H2O 2Pi, Go’ = -29 kJ mol-1 • Rapidity of this hydrolysis drives reactions involving pyrophosphate to right Metabolism; Glycolysis
ATP chemistry • Why is ATP a high-energy compound? • Negative charges repel!(somewhat mitigated by Mg2+) • ADP and Pi or AMP and PPi are better solvated than ATP • More delocalization in products • Therefore ATP (and CTP, GTP, …)are high-energy compounds Metabolism; Glycolysis
Interconversions among nucleotide phosphates • Kinases or phospotransferases involved • Nucleoside monophosphate kinases:ATP + XMP ADP + XDPspecific to each X (G or dG, C or dC, …) • Nucleoside diphosphate kinase:ATP + XDP ADP + XTPthis is a single enzyme (why?) Metabolism; Glycolysis
AnP interconversions • Adenylate kinase (special case of NMK…) • AMP + ATP 2ADP • [ATP] >> [ADP] and [ATP] >> [AMP], so small changes in [ATP] can drive big changes in others: [ATP],mM [ADP],mM [AMP],mM G1,kJmol-1 4.8 0.2 0.004 -40 4.5 0.5 0.02 -37 3.9 1.0 0.11 -35 3.2 1.5 0.31 -34 Metabolism; Glycolysis
How coupling works • We tend to hand-wave about this • Enzymes provide location for intermediates • X + ATP X—P + ADPX—P + Y + H2O X-Y + Pi + H+ • The X here can be an enzyme side-chain or a substrate • In the former case some other event must come along to recreate X;otherwise, it isn’t an enzyme! Metabolism; Glycolysis
Glutamine synthetase • Cf. section 25.2 - 25.3: • glu + ATP -glutamylphosphate + ADP • -glutamylphosphate + NH3 -> gln + Pi • Why do we need ATP at all for this? • Go’ = 14 kJ mol-1 for glu + NH3 -> gln + H2O • we could overcome that with concentrations • But we can’t: we need [glu] ~ [gln] • So we need the energy charge Metabolism; Glycolysis
CompoundGo’hyd, kJ mol-1 PEP -62 1,3-bisPglycerate -49 ATP->AMP+PPi -45 Phosphocreatine -43 P-arginine -32 CompoundGo’hyd Acetyl CoA* -32 ATP -32 Pyrophosphate -29 Glucose 1-P -21 Glucose 6-P -14 Glycerol 3-P -9* not a phosphate cmpd! Go’hyd for metabolites Transfers are more common than hydrolysis; these help with bookkeeping Metabolism; Glycolysis
Making ATP by transfer • We’ve just seen that some compounds have higher-energy phosphates than ATP • Remember the 35-cent analogy • So a phosphoryl-group transfer into ATP can be energetically favorable, e.g. Phosphocreatine + ADP creatine + ATP Metabolism; Glycolysis
Relative energies of phosphate compounds • Both phosphoenolpyruvate(~ 60 kJ mol-1) and phosphocreatine(~ 45 kJ mol-1) are higher-energy than ATP (*what does that mean?) • ATP is therefore intermediate between these high-energy phosphates and the low-energy phosphates like glucose-6-phosphate and glucose-1-phosphate Metabolism; Glycolysis
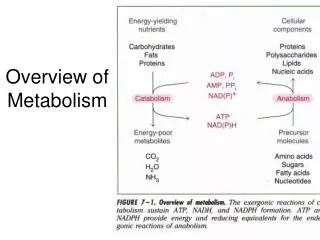
ATP and the energy cycle • Catabolism usually gives rise to energy that is captured in high-energy phosphate bonds in ATP • This ATP is used to provide energy for otherwise endergonic reactions and to phosphorylate things that need to be phosphorylated Metabolism; Glycolysis
Nucleotidyl-group transfer • It’s most convenient to think of this as a transfer of the entire nucleotide group to form an acyl-adenylate intermediate • This can then fall apart, releasing AMP and allowing a high-energy bond to form • Example: acetyl CoA (see pp.561-562) Metabolism; Glycolysis
Thioesters: another class of high-energy compounds • Thioesters have similar reactivity as oxygen-acid anhydrides • Thioesters less stable than oxygen esters because the unshared electrons in sulfur are not as delocalized in a thioester as the unshared electrons in an oxygen ester Metabolism; Glycolysis
Oxidation-reduction reactions and Energy • Oxidation-reduction reactions involve transfer of electrons, often along with other things • Generally compounds with many C-H bonds are high in energy because the carbons can be oxidized (can lose electrons) Metabolism; Glycolysis
Reduction potential • Reduction potential is a measure of thermodynamic activity in the context of movement of electrons • Described in terms of half-reactions • Each half-reaction has an electrical potential, measured in volts, associated with it because we can (in principle) measure it in an electrochemical cell Metabolism; Glycolysis
So what is voltage, anyway? • Electrical potential is available energy per unit charge: • 1 volt = 1 Joule per coulomb • 1 coulomb = 6.24*1018 electrons • Therefore energy is equal to the potential multiplied by the number of electrons Metabolism; Glycolysis
Electrical potential and energy • This can be expressed thus:Go’ = -nFEo’ • n is the number of electrons transferred • F = fancy way of writing # of Coulombs (which is how we measure charge) in a mole (which is how we calibrate our energies) = 96.48 kJ V-1mol-1 Metabolism; Glycolysis
Oh yeah? • Yes. • 1 mole of electrons = 6.022 * 1023 e- • 1 coulomb = 6.24*1018 e- • 1 mole = 9.648*104 Coulomb • 1 V = 1 J / Coulomb=10-3 kJ / Coulomb • Therefore the energy per mole associated with one volt is10-3 kJ / C * 9.648*104 C = 96.48 kJ Metabolism; Glycolysis
What can we do with that? • The relevant voltage is the difference in standard reduction potential between two half-reactions • Eo’ = Eo’acceptor - Eo’donor • Combined with free energy calc, we seeEo’ = (RT/nF ) lnKeq andE = Eo’ - (RT/nF ) ln [products]/[reactants] • This is the Nernst equation Metabolism; Glycolysis
Free energy from electron transfer • We can examine tables of electrochemical half-reactions to get an idea of the yield or requirement for energy in redox reactions • Example (see section 10.9B):NADH + (1/2)O2 + H+ -> NAD+ + H2O; • We can break that up into half-reactions to determine the energies Metabolism; Glycolysis
Half-reactions and energy • NAD+ + 2H+ + 2e- NADH + H+,Eo’ = -0.32V • (1/2)O2 + 2H+ + 2e- H2O, Eo’ = 0.82V • Reverse the first reaction and add:NADH + (1/2)O2 + H+ NAD+ + H2O,Eo’ = 0.82+0.32V = 1.14 V. • Go’ = -nFEo’ = -2*(96.48 kJ V-1mol-1)(1.14V) = -220 kJ mol-1; that’s a lot! Metabolism; Glycolysis
NAD: electron collector • Net reactions involve transfer of hydride (H:-) ions • Enzymes called dehydrogenases (a type of oxidoreductase) involved • Collected NADH can then be reoxidized in oxidative phosphorylation to drive ATP synthesis in the mitochondrion Metabolism; Glycolysis
NADPH • Provides reducing power for anabolic reactions • Often converting highly oxidized sugar precursors into less reduced molecules Metabolism; Glycolysis
NAD+ 340 nm How to detect NAD reactions Absorbance NADH Wavelength • NAD+ and NADH(and NADP+ and NADPH)have extended aromatic systems • But the nicotinamide ring absorbs strongly at 340 only in the reduced(NADH, NADPH) forms • Spectrum is almost pH-independent, too! • So we can monitor NAD and NADP-dependent reactions by appearance or disappearance of absorption at 340 nm Metabolism; Glycolysis
How much ATP can we get out of oxidizing NADH? • In principle -220 kJ mol-1 should be enough to drive production of at least five ATP molecules • (220/32) = 6.9; even if we figure it will cost more like 40 kJ mol-1 per ATP, then that’s (220/40) = 5.5. • But in fact we only get about 3.5. Metabolism; Glycolysis
Why? • Short answer: discrete inefficiency • 4 oxidation steps in the electron transport chain beginning with NADH • 3 of 4 of those steps facilitate transfer of protons against a pH and charge gradient • When those protons move back across with their charge and concentration gradients, we earn ATP back • … but only about net 3.5 ATP per NADH Metabolism; Glycolysis
Pathway methods • Introducing inhibitors • Site-directed mutagenesis • Radioisotope tracing • Non-radioactive isotope tracing • NMR Metabolism; Glycolysis
Classical metabolism studies • Add substrate to a prep and look for intermediates and end products • If substrate is radiolabeled (3H, 14C) it’s easier, but even nonradioactive isotopes can be used for mass spectrometry and NMR • NMR on protons, 13C, 15N, 31P • Reproduce reactions using isolated substrates and enzymes Metabolism; Glycolysis
Next level of sophistication… • Look at metabolite concentrations in intact cell or organism under relevant physiological conditions • Note that Km is often ~ [S].If that isn’t true, maybe you’re looking at the non-physiological substrate! • Think about what’s really present in the cell. Metabolism; Glycolysis
Mutations in single genes • If we observe or create a mutation in a single gene of an organism, we can find out what the effects on viability and metabolism are • In humans we can observe genetic diseases and tease out the defective gene and its protein or tRNA product • Sometimes there are compensating enzyme systems that take over when one enzyme is dead or operating incorrectly Metabolism; Glycolysis
Deliberate manipulations • Bacteria and yeast: • Irradiation or exposure to chemical mutagens • Site-directed mutagenesis • Higher organisms:We can delete or nullify some genes;thus knockout mice • Introduce inhibitors to pathways and see what accumulates and what fails to be synthesized Metabolism; Glycolysis
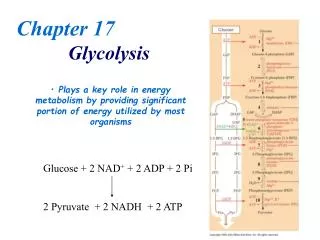
Glycolysis • Now we’re ready for the specifics of metabolism • Why glycolysis first? • Well-understood (?) early on • Illustrates concepts used later • Inherently important Metabolism; Glycolysis
The big picture • Conversion of glucose to pyruvate • Catabolic, ten steps, energy-yielding • Overall reaction: glucose + 2 ADP + 2 NAD+ + 2Pi2pyruvate + 2ATP + 2NADH + 2H+ + 2H2O Metabolism; Glycolysis
Significance • Why is this important? • Energy production(ATP and NADH) • Pyruvate as precursor to various metabolites • Some steps require energy • So it isn’t all energy-yielding • The net reaction yields energy Metabolism; Glycolysis
The reactions • See fig. 11.2 and the table in the HTML notes • Wide variety of enzyme sizes • All then enzyme structures have been determined by X-ray crystallography, mostly at high resolution Metabolism; Glycolysis
The pathway through TIM Figure Courtesy U.Texas Metabolism; Glycolysis
Pathway: TIM to pyruvate Bottom half of same graphic Metabolism; Glycolysis