Download
1 / 19
190 likes | 333 Views
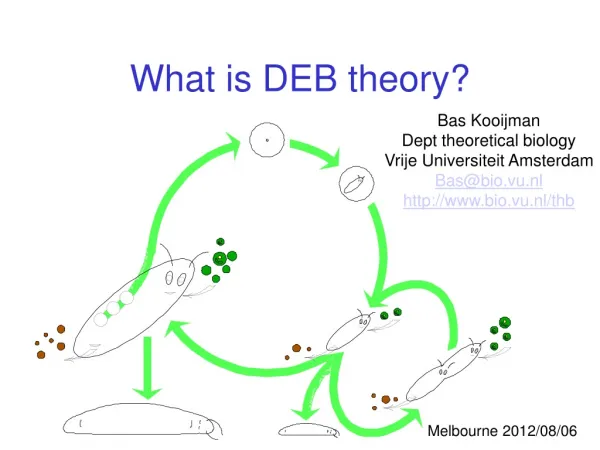
Applications of DEB theory. Bas Kooijman Dept theoretical biology Vrije Universiteit Amsterdam Bas@bio.vu.nl http://www.bio.vu.nl/thb. Iraklion, 2010/05/12. Contents. Very short outline of standard DEB model Reconstruction of food/temperature trajectories

E N D
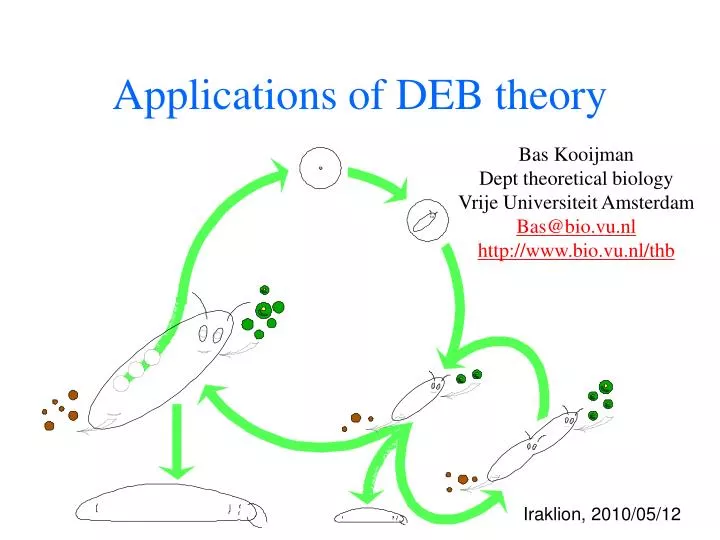
Applications of DEB theory Bas Kooijman Dept theoretical biology Vrije Universiteit Amsterdam Bas@bio.vu.nl http://www.bio.vu.nl/thb Iraklion, 2010/05/12
Contents • Very short outline of standard DEB model • Reconstruction of food/temperature trajectories • Identification of mode of action of toxicants • Prediction of tumour growth as affected by caloric restriction • Optimisation of bioproduction yield of microbial biomass prediction of spawning by pacific oysters
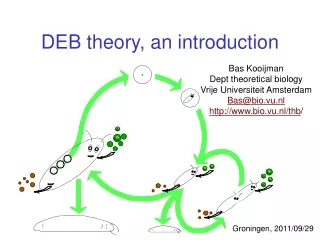
defecation feeding food faeces assimilation reserve somatic maintenance maturity maintenance 1- maturation reproduction growth maturity offspring structure Standard DEB model 1 food type, 1 reserve, 1 structure, isomorph time: searching & handling feeding surface area weak & strong homeostasis κ-rule for allocation to soma maintenance has priority somatic maint structure maturity maint maturity stage transition: maturation embryo: no feeding, reprod juvenile: no reproduction adult: no maturation maternal effect: reserve density at birth equals that of mother initially: zero structure, maturity
Food intake from weights Aptenodytes forsteri Pygoscelis adelidae food intake, cm2 weight1/3, g1/3 weight1/3, g1/3 age, d age, d Small-bodied pinguins synchronise breeding with food peak and grow von Bertalanffy Large-bodied pinguins: fit spline through weights reconstruct food intake, given DEB parameters Kooijman 1993 Cambridge Univ Press
Arrhenius relationship Arrhenius ln pop growth rate, h-1 Escherichia coli 103/T, K-1 Kooijman 2000 Cambridge Univ Press 103/TH 103/TL
Body temperature from weights Uria aalge Mahoney & Trelfall 1981 Catharacta skua Furness 1987 weight1/3, g1/3 weight1/3, g1/3 body temperature, °C body temperature, °C age, d age, d fit generalised logistic growth through weights assume abudant food reconstruct body temperature, given DEB parameters and Arrhenius relationship Kooijman 1993 Cambridge Univ Press
Food density from reproduction • Eggs in brood pouches of Daphnia hyalina were counted • in weekly hauls in 4 mesocosms by Stella Berger (München) • and eggs and body length are measured • Given DEB parameters for D. hyalina • scaled functional response can be estimated as functions of time • based on assumptions: • functional response is constant during a week • and individuals are in pseudo-equilibrium • all individuals have equal parameters but experience different food densities • (estimate is individual-specific for each time point) • or • individuals differ in parameters but experience the same food density • DEB elements: • handling rules for reproduction buffer comply to the molting cycle • maternal effect: reserve density at birth equals that of the mother at egg formation • eggs grow in volume during incubation due to uptake of water Kooijman 2010 Cambridge Univ Press
Food density from reproduction food different for each ind food the same for ind in one haul initial egg volume, mm3 f f scaled functional resp, f week week
Analysis of otolith data North Sea cod Barents Sea cod observed simulated Otolith formation: organic matrix (OM) has contributions from dissipation and growth CaCO3 is stoichiometrically coupled in a temperature-dependent way opacity reflects the CaCO3-OM ratio Fablet et al 2010, in prep Gadus morhua
Analysis of otolith data † May 1 April 15 checks simulated scenario’s Fablet et al 2010, in prep
Food & temperature reconstruction from otolith data fitted observed fitted if food constant Direct observation: feeding was low for low and high temperatures Fablet et al 2010, in prep
TPT body length cumulative offspring time time Increase in maintenance costs Folsomia candida Tri-Phenyl-Tin Jager et al (2004) Environ Sci Technol 38: 2894-2900
Chlorpyrifos body length cumulative offspring time time Increase in cost for offspring Folsomia candida Jager et al (2007) Environ Pollut 145: 452-458
Pentachlorobenzene body length cumulative offspring time time Increase in cost for structure Caenorhabditis elegans Alda Álvarez et al (2006) Environ Sci Technol 40:2478-2484
Cd & Cu survival of Folsomia Interaction Cu,Cd, Pb, Zn: Cu & Pb: slightly antagonistic Other combinations: nill Folsomia candida Baas et al 2007 Eviron Tox Chem26: 1320-1327
Tumour Growth: workload allocation Growth curve of tumour depends on pars no maximum size is assumed a priori If tumour induction occurs late, tumours grows slower Van Leeuwen et al 2003 Brit J Cancer89: 2254-2263
Yield vs growth Streptococcus bovis, Russell & Baldwin (1979) Marr-Pirt (no reserve) DEB 1/yield, mmol glucose/ mg cells spec growth rate yield 1/spec growth rate, h Russell & Cook (1995): this is evidence for down-regulation of maintenance at high growth rates DEB theory: high reserve density gives high growth rates structure requires maintenance, reserves do not Kooijman & Troost 2007 Biol Rev 82: 1-30
Growth & reprod in Crassostrea gigas • Bay of Arcachon • DEB curves based on • measured temperature • measured chlorophyll • known DEB parameters • Spawings (jumps in weight) • correctly predicted • on the basis of temp trigger Pouvreau et al 2006 J Sea Res56: 156-167
DEB tele course 2011 http://www.bio.vu.nl/thb/deb/ Course is free of financial costs; some 100 h effort Program for 2011: Feb/Mar general theory (5w) 13-15 April course +symposium in Lisbon (2w + 3 d) Target audience: PhD students We encourage participation in groups who organize local meetings weekly Software package DEBtool for Octave/ Matlab freely downloadable Slides of this presentation are downloadable from http://www.bio.vu.nl/thb/users/bas/lectures/ Cambridge Univ Press 2009 Audience: thank you for your attention