Download

1 / 27
330 likes | 1.03k Views
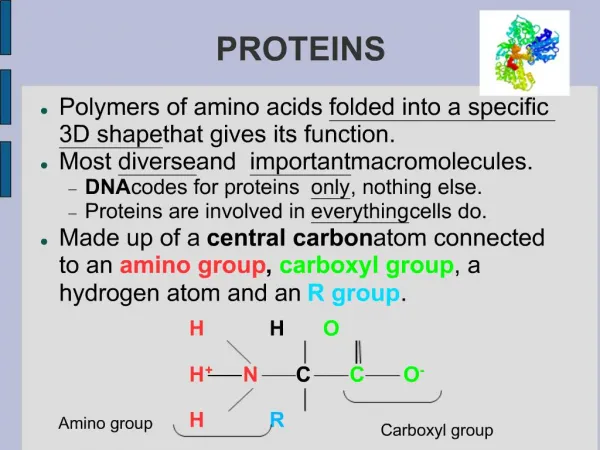
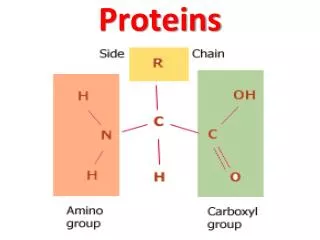
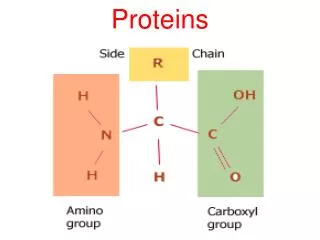
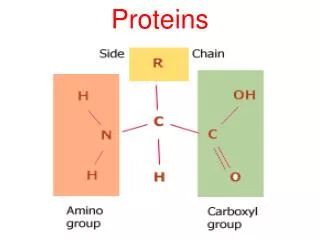
MUSCLE PROTEINS. What is the mechanism whereby conformational changes in biological molecules is translated into macroscopic movements?. Skeletal Muscle Anatomy. A fiber bundle contains hundreds of myofibrils that run the length of the fiber Each myofibril is a linear array of sarcomeres

E N D
MUSCLE PROTEINS • What is the mechanism whereby conformational changes in biological molecules is translated into macroscopic movements?
Skeletal Muscle Anatomy • A fiber bundle contains hundreds of myofibrils that run the length of the fiber • Each myofibril is a linear array of sarcomeres • Each sarcomere is capped on ends by a transverse tubule (t-tubule) that is an extension of sarcolemmal membrane • Surfaces of sarcomeres are covered by sarcoplasmic reticulum containing calcium
16.1 What Is a Molecular Motor? Figure 16.1 The structure of a skeletal muscle cell, showing the manner in which transverse tubules enable the sarcolemmal membrane to extend into the interior of the fiber.
Electron micrograph of a skeletal muscle myofibril. The length of one sarcomere is indicated, as are the A and I bands, the H zone, the M disk, and the Z lines. Cross-sections from the H zone show a hexagonal array of thick filaments, whereas the I band cross-section shows a hexagonal array of thin filaments. The major chemical entities here are F-actin and myosin.
The three-dimensional structure of an actinmonomer from skeletal muscle. This view shows the two domains (left and right) of actin. This component in fibrillar form, F-actin is the material in the thin-fillament potion of the sarcomere.
The helical arrangement of actin monomers in F-actin. The F-actin helix has a pitch of 72 nm and a repeat distance of 36 nm.
(a) An electron micrograph of a thinfilament, (b) a corresponding image reconstruction, and (c) a schematic drawing based on the images in (a) and (b). The tropomyosin coiled coil winds around the actin helix, each tropomyosin dimer interacting with seven consecutive actin monomers. Troponin T binds to tropomyosin
The Composition and Structure of Thick Filaments Myosin - 2 heavy chains, 4 light chains • Heavy chains - 230 kD each • Light chains - 2 pairs of different 20 kD chains • The "heads" of heavy chains have ATPase activity and hydrolysis here drives contraction • Light chains regulate movement
An electron micrograph of a myosin molecule and a corresponding schematic drawing. The tail is a coiled coil of intertwined a-helices extending from the two globular heads. One of each of the myosin light chain proteins, LC1 and LC2, is bound to each of the globular heads. Loss of the LC1 chain abolishes ATPase activity of the myosin heads. • A ribbon diagram shows the structure of the S1 myosin head (green, red, and purple segments) and its associated essential (yellow) and regulatory (magenta) light chains.
Repeating Structural Elements Are the Secret of Myosin’s Coiled Coils The secret to ultrastructure • 7-residue, 28-residue and 196-residue repeats are responsible for the organization of thick filaments • Residues 1 and 4 (a and d) of the seven-residue repeat are hydrophobic; residues 2,3 and 6 (b, c and f) are ionic • This repeating pattern favors formation of coiled coil of tails.
Figure 16.17An axial view of the two-stranded, a-helical coiled coil of a myosin tail. Hydrophobic residues a and d (1 and 4)of the seven-residue repeat sequence align to form a hydrophobic core. Residues b, c, and f (2,3 and 6) face the outer surface of the coiled coil and are typically ionic.
More Repeats! • 28-residue repeat (4 x 7) consists of distinct patterns of alternating side-chain charge (+ vs -), and these regions pack with regions of opposite charge on adjacent myosins to stabilize the filament • 196-residue repeat (7 x 28) pattern also contributes to packing and stability of filaments • Indeed, myosin molecules in the filament are staggered relative to one another by 98 residues, one-half the residue repeat
The packing of myosin molecules in a thick filament. Adjoining molecules are offset by approximately 14 nm, a distance corresponding to 98 residues of the coiled coil.
Electron micrograph of a skeletal muscle myofibril. The length of one sarcomere is indicated, as are the A and I bands, the H zone, the M disk, and the Z lines. Cross-sections from the H zone show a hexagonal array of thick filaments, whereas the I band cross-section shows a hexagonal array of thin filaments. The major chemical entities here are F-actin and myosin.
How does the system work? The sliding filament model of skeletal muscle contraction. The decrease in sarcomere length is due to decreases in the width of the Iband and H zone, with no change in the width of the A band. These observations mean that the lengths of both the thick and thin filaments do not change during contraction. Rather, the thick and thin filaments slide along one another.
The Contraction Cycle: ATP Hydrolysis Drives Conformation Changes in Myosin Figure 16.9 The mechanism of skeletal muscle contraction. The free energy of ATP hydrolysis drives a conformational change in the myosin head, resulting in net movement of the myosin heads along the actin filament.
The Contraction Cycle: ATP Hydrolysis Drives Conformation Changes in Myosin Cross-bridge formation is followed by power stroke with ADP and Pi release ATP binding causes dissociation of myosin heads and reorientation of myosin head Details of the conformational change in the myosin heads are coming to light Evidence now exists for a movement of at least 35 Å in the conformation change between the ADP-bound state and ADP-free state
Ca2+ is the trigger signal for muscle contraction. Release of Ca2+ through voltage- or Ca2+-sensitive channels activates contraction. Ca2+ pumps induce relaxation by reducing the concentration of Ca2+ available to the muscle fibers.
A drawing of the thick and thin filaments of skeletal muscle in cross-section showing the changes that are postulated to occur when Ca2+ binds to troponin C.
Smooth Muscle Contraction No troponin complex in smooth muscle • In smooth muscle, Ca2+ activates myosin light chain kinase (MLCK) which phosphorylates LC2, the regulatory light chain of myosin • Ca2+ effect is via calmodulin - a cousin of TnC • Hormones regulate contraction - epinephrine, a smooth muscle relaxer, activates adenylyl cyclase, making cAMP, which activates protein kinase, which phosphorylates MLCK, inactivating MLCK and relaxing muscle
Smooth Muscle Effectors Useful drugs • Epinephrine (as Primatene) is an over-the-counter asthma drug, but it acts on heart as well as on lungs - a possible problem! • Albuterol is a more selective smooth muscle relaxer and acts more on lungs than heart • Albuterol is used to prevent premature labor • Oxytocin (pitocin) stimulates contraction of uterine smooth muscle, inducing labor
Kinases and Phosphatases • Kinases add a phosphate group to another molecule from a donor such as ATP • Phosphatases remove phosphate groups • On proteins the presence of a phosphate group on a serine/threonine/tyrosine –OH can have regulatory implications (e.g. on myosin light chains in smooth muscle).
Creatine Kinase and Phosphocreatine Provide an Energy Reserve in Muscle Figure 27.9 Phosphocreatine serves as a reservoir of ATP-synthesizing potential.
Lactate Formed in Muscles is Recycled to Glucose in the Liver How your liver helps you during exercise: Recall that vigorous exercise can lead to a buildup of lactate and NADH, due to oxygen shortage and the need for more glycolysis NADH can be reoxidized during the reduction of pyruvate to lactate Lactate is then returned to the liver, where it can be reoxidized to pyruvate by liver LDH Liver provides glucose to muscle for exercise and then reprocesses lactate into new glucose This is referred to as the Cori cycle (Figure 22.7)
Lactate Formed in Muscles is Recycled to Glucose in the Liver Figure 22.7 The Cori cycle.
Muscle control by receptor agonism and antagonism • Tropicamide, an antimuscarinic drug causes dilation of the pupil by inhibiting the Acetylcholine receptors of the circular muscles in the iris. Sympathomimetics such as phenylephrine HCl are used to directly stimulate the radial muscles of iris.