Download

1 / 21
270 likes | 694 Views
Enzymes. Introduction. Headlines of the day…. TRPM2: a multifunctional ion channel for calcium signalling Published online before print December 6, 2010, doi : 10.1113/jphysiol.2010.201855 J Physiol April 1, 2011 vol. 589 no. 7 1515-1525 .

E N D
Enzymes Introduction
TRPM2: a multifunctional ion channel for calcium signalling Published online before print December 6, 2010, doi: 10.1113/jphysiol.2010.201855 J Physiol April 1, 2011 vol. 589 no. 7 1515-1525 Figure 2. Signalling mechanisms for TRPM2 activation NAD+ and reactive oxygen species (ROS), including H2O2, accumulate during inflammation and tissue damage. External NAD+ may be converted to ADPR, cADPR and NAADP by the ectoenzymes CD38 and CD157. Extracellular ADPR may then bind to plasma membrane receptors (e.g. P2Y receptors) and increases [Ca2+]i through Ca2+ release from stores via G-proteins and phospholipase C (PLC) activation with subsequent IP3 production. H2O2 may also cross the plasma membrane and mobilize ADPR from mitochondria (both H2O2 and cADPR can synergize with ADPR to activate TRPM2). ADPR is also generated from poly-ADPR during ROS-induced DNA damage through activation of the PARP-1/PARG pathway. Free cytosolic ADPR will act on the NUDT9-H of lysosomal and plasma membrane TRPM2 channels, enabling Ca2+ influx across the plasma membrane and/or release of lysosomal Ca2+, raising the Ca2+ concentration in the cytosol. Ca2+ overload can trigger programmed cell death (apoptosis) and possibly necrosis. Finally, extracellular signals that remain to be identified could potentially induce the production of intracellular free ADPR, which may then gate TRPM2 channels in the lysosome and/or plasma membrane and regulate receptor-mediated signalling.
Headlines of the day • The membrane-bound enzyme CD38 exists in two opposing orientations. • Zhao YJ, Lam CM, Lee HC. • Source:Department of Physiology, Li Ka Shing School of Medicine, The University of Hong Kong, Hong Kong, China. • Abstract • The transmembrane enzyme CD38, a multifunctional protein ubiquitously present in cells, is the main enzyme that synthesizes and hydrolyzes cyclic adenosine 5'-diphosphate-ribose (cADPR), an intracellular Ca(2+)-mobilizing messenger. CD38 is thought to be a type II transmembrane protein with its carboxyl-terminal catalytic domain located on the outside of the cell; thus, the mechanism by which CD38 metabolizes intracellular cADPR has been controversial. We developed specific antibodies against the amino-terminal segment of CD38 and showed that two opposing orientations of CD38, type II and type III (which has its catalytic domain inside the cell), were both present on the surface of HL-60 cells during retinoic acid-induced differentiation. When activated by interferon-γ, human primary monocytes and the monocytic U937 cell line exhibited a similar co-distribution pattern. Site-directed mutagenesis experiments showed that the membrane orientation of CD38 could be converted from a mixture of type II and type III orientations to all type III by mutating the cationic amino acid residues in the amino-terminal segment of CD38. Expression of type III CD38 construct in transfected cells led to increased intracellular concentrations of cADPR, indicating the importance of the type III orientation of CD38 to its Ca(2+) signaling function. The identification of these two forms of CD38 suggests that flipping the catalytic domain from the outside to the inside of the cell may be a mechanism regulating its signaling activity.
GENERAL CHARACTERISTICS • Most enzymes are protein, of the globular form. • Being proteins, they are affected by extremes of heat, pH and radiation. • All enzymes speed up the rate of a reaction. • All enzyme catalyzed reactions are reversible. • Enzymes speed up the rate of reaction by lowering the energy of activation, i.e. Ea. • Enzymes are not involved in the reaction. • They are not destroyed at the end of a reaction. • They are either intracellular or extracellular in function. • Enzymes work only in solution. • All enzyme catalyzed reactions are affected by substrate concentration and enzyme concentration. • All enzymes are specific for their substrate.
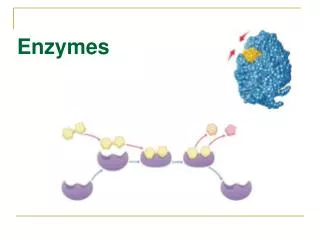
How enzymes work? • Enzymes attach only to their specific substrate. • They remain in an enzyme-substrate complex. • They breakdown/build the substrate into respective products. • Enzyme dissociates itself from the product. E+S=ES complex=E+P
Activation energy The energy that an atomic system must acquire before a process (such as an emission or reaction) can occur; "catalysts are said to reduce the energy of activation during the transition phase of a reaction."
Structure and mechanism Structures and mechanisms Enzymes are generally globular proteins and range from 62 amino acid residues in size, to over 2,500 residues in the animal fatty acid synthase. The activities of enzymes are determined by their three-dimensional structure Most enzymes are much larger than the substrates they act on, and only a small portion of the enzyme (around 3–4 amino acids) is directly involved in catalysis. Giorgio Lampis, Alessandra Desogus, Sabrina Petruzzelli, SamuelaLaconi, Angela Ingianni, Maria AntoniettaMadeddu, RaffaelloPompe, Anaerobe, Volume 13, Issues 5–6, October–December 2007, Pages 238–243
Dalton…unit of mass The dalton, symbol Da, is also sometimes used as a unit of molar mass, especially in biochemistry, with the definition 1 Da = 1 g/mol, despite the fact that it is strictly a unit of mass (1 Da = 1.660 538 782(83)×10−27 kg.)
Maltase enzyme Maltose is made of two glucose molecules bonded together (1). The maltase enzyme is a protein that is perfectly shaped to accept a maltose molecule and break the bond (2). The two glucose molecules are released (3). A single maltase enzyme can break in excess of 1,000 maltose bonds per second, and will only accept maltose molecules. Characteristics of Maltase from Baker's Yeast: Molecular weight: 68,500 daltons. pH Optimum: 7.0 - 7.5 using maltose as the substrate. Inhibitors: Thiol blocking compounds, heavy metal ions, histidine, and certain amines. Tris should not be used as a buffer due to its inhibitory effect. Storage: Store at 2-8° C.
Active site The region that contains the catalytic residues, binds the substrate, and then carries out the reaction is known as the active site. Enzymes can also contain sites that bind cofactors, which are needed for catalysis. Some enzymes also have binding sites for small molecules, which are often direct or indirect products or substrates of the reaction catalyzed. This binding can serve to increase or decrease the enzyme's activity, providing a means for feedback regulation.
Jesse saying no to Bessie…politely The frequency of decreased lactase activity ranges from 5% in northern Europe through 71% for Sicily to more than 90% in some African and Asian countries.
Denaturation and reversibility Most enzymes can be denatured—that is, unfolded and inactivated—by heating, which destroys the three-dimensional structure of the protein. Depending on the enzyme, denaturation may be reversible or irreversible.
Models of enzyme action "Lock and key" model : "Lock and key" model Enzymes are very specific, because both the enzyme and the substrate possess specific complementary geometric shapes that fit exactly into one another. This is often referred to as "the lock and key" model. However, while this model explains enzyme specificity, it fails to explain the stabilization of the transition state that enzymes achieve. Induced fit model : Induced fit model Diagrams to show the induced fit hypothesis of enzyme action. since enzymes are rather flexible structures, the active site is continually reshaped by interactions with the substrate as the substrate interacts with the enzyme. As a result, the substrate does not simply bind to a rigid active site. In some cases, such as glycosidases , the substrate molecule also changes shape slightly as it enters the active site.
Transition State Model • Transition State Theory: • In the transition state theory, the mechanism of interaction of reactants is not considered; the important criterion is that colliding molecules must have sufficient energy to overcome a potential energy barrier (the activation energy) to react. • It takes a lot of energy to achieve the transition state, so the state is a high-energy substance. The potential energy of the system increases at this point because: • The approaching reactant molecules must overcome the mutual repulsive forces between the outer shell electrons of their constituent atoms. • Atoms must be separated from each other as bonds are broken.
Lock and Key Model + + E + S ES complex E + P P S S P
Enzyme Action: Lock and Key Model • An enzyme binds a substrate in a region called the active site • Only certain substrates can fit the active site • Amino acid R groups in the active site help substrate bind • Enzyme-substrate complex forms • Substrate reacts to form product • Product is released
Enzyme Action: Induced Fit Model • E + S ES complex E + P P S S S P
Enzyme Action: Induced Fit Model • Enzyme structure flexible, not rigid • Enzyme and active site adjust shape to bind substrate • Increases range of substrate specificity • Shape changes also improve catalysis during reaction
Transition State Model For a reaction involving two molecules, a transition state is formed when the �old bonds between two molecules are weakened and new bonds begin to form or the old bonds break first to form the transition state and then the new bonds form after. The theory suggests that as reactant molecules approach each other closely they are momentarily in a less stable state than either the reactants or the products. In the example below, the first scenario occurs to form the transition state: