Download

1 / 53
560 likes | 825 Views
Muscle contraction. The term " excitation-contraction coupling " refers to the mechanism by which the action potential causes the myofibrils of muscle to contract. Each muscle fiber behaves as a single unit, is multinucleate, and contains myofibrils. The myofibrils are surrounded by

E N D
The term "excitation-contraction coupling" refers to the mechanism by which the action potential causes the myofibrils of muscle to contract
Each muscle fiber behaves as • a single unit, is multinucleate, • and contains myofibrils. • The myofibrils are surrounded by • sarcoplasmic reticulum and are • invaginated by • transverse tubules (T tubules). • Each myofibril contains interdigitating thick and thin filaments, which are arranged longitudinally and cross-sectionally in sarcomeres • The repeating units of sarcomeres account for the unique banding pattern seen in striated muscle (which includes both skeletal and cardiac muscle
surrounding the myofibrils of each muscle fiber is an extensive reticulum called thesarcoplasmic reticulum. • This reticulum has a special organization that is extremely important in controlling muscle contraction.
Transverse Tubules and the Sarcoplasmic Reticulum Thetransverse (T) tubulesare an extensive network of muscle cell membrane (sarcolemmal membrane) that invaginates deep into the muscle fiber. The T tubules are responsible for carrying depolarization from action potentials at the muscle cell surface to the interior of the fiber. Thesarcoplasmic reticulumis an internal tubular structure, which is the site of storage and release of Ca2+ for excitation-contraction coupling.
the myosin heads have an actin-binding site, which is necessary for cross-bridge formation, and a site that binds and hydrolyzes ATP (myosin ATPase( • Actin has myosin-binding sites. When the muscle is at rest, the myosin-binding sites are covered by tropomyosin so that actin and myosin cannot interact. • If contraction is to occur, tropomyosin must be moved out of the way so that actin and myosin can interact.
the tropomyosin molecules lie on top of the active sites of the actin strands so that attraction cannot occur between the actin and myosin filaments to cause contraction. • When calcium ions combine with troponin C, the troponin complex supposedly undergoes a conformational change that in some way "uncovers" the active sites of the actin, thus allowing these to attract the myosin cross-bridge heads and cause contraction to proceed.
It is the interaction between these cross-bridges of myocin and the actin filaments that causes contraction.
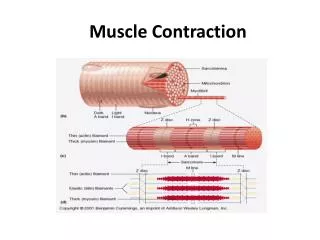
Arrangement of Thick and Thin Filaments in Sarcomeres Thesarcomereis the basic contractile unit, and it is delineated by: the Z disks. Each sarcomere contains a full A band in the center and one half of two I bands on either side of the A band
at sarcomere length of about 2 micrometers the muscle is capable of generating its greatest force of contraction.
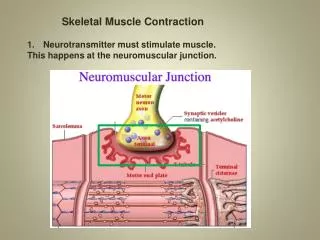
The initiation and execution of muscle contraction occur in the following sequential steps. 1- An action potential travels along a motor nerve to its endings on muscle fibers 2- At each ending, the nerve secretes a small amount of the neurotransmitter substanceacetylcholine. 3- The acetylcholine acts on a local area of the muscle fiber membrane to open multiple "acetylcholine-gated" cation channels through protein molecules floating in the membrane
4- Opening of the acetylcholine-gated channels allows large quantities of sodium ions to diffuse to the interior of the muscle fiber membrane. This causes a local depolarization that in turn leads to opening of voltage-gated sodium channels. This initiates an action potential at the membrane. 5- The action potential travels along the muscle fiber membrane in the same way that action potentials travel along nerve fiber membranes 6- The action potential depolarizes the muscle membrane, and much of the action potential electricity flows through the center of the muscle fiber. Here it causes the sarcoplasmic reticulum to release large quantities of calcium ions that have been stored within this reticulum.
7- The calcium ions initiate attractive forces between the actin and myosin filaments, causing them to slide alongside each other, which is the contractile process 8- After a fraction of a second, the calcium ions are pumped back into the sarcoplasmic reticulum by a Ca++ membrane pump and remain stored in the reticulum until a new muscle action potential comes along; this removal of calcium ions from the myofibrils causes the muscle contraction to cease.
EXCITATION-CONTRACTION COUPLING IN SKELETAL MUSCLE • shows the temporal relationships between an action potential in the skeletal muscle fiber, the subsequent increase in intracellular free Ca2+ concentration and contraction of the muscle fiber. • These temporal relationships are critical in that the action potential alwaysprecedesthe rise in intracellular Ca2+ concentration, which alwaysprecedescontraction.
muscle contraction occurs by asliding filament mechanism This is caused by forces generated by interaction of the cross-bridges from the myosin filaments with the actin filaments.
Interaction Between the "Activated" Actin Filament and the Myosin Cross-Bridges- The "Walk-Along" Theory of Contraction The head attaches to an active site, this causes forces between the head and arm of its cross-bridge. this causes the head to tilt toward the arm and to drag the actin filament along with it. This tilt of the head is called thepower stroke. Then, immediately after tilting, the head automatically breaks away from the active site. Next, the head returns to its extended direction. In this position, it combines with a new active site farther down along the actin filament; then the head tilts again to cause a new power stroke, and the actin filament moves another step
Thus, the heads of the cross-bridges bend back and forth and step by step walk along the actin filament, pulling the ends of two successive actin filaments toward the center of the myosin filament
A,At the beginning of the cycle, no ATP is bound to myosin, and myosin is tightly attached to actin in a "rigor" position. In rapidly contracting muscle, this state is very brief. However, in the absence of ATP, this state is permanent (i.e., rigor mortis
B,The binding of ATP to a cleft on the back of the myosin head produces a conformational change in myosin that decreases its affinity for actin; thus, myosin is released from the original actin-binding site. C,
C,The cleft closes around the bound ATP molecule, producing a further conformational change that causes myosin to be displaced toward the plus end of actin. ATP is hydrolyzed to ADP and Pi, which remain attached to myosin
D,Myosin binds to a new site on actin (toward the plus end), constituting the force-generating, or power, stroke. Each cross-bridge cycle "walks" the myosin head 10 nanometers along the actin filament
E,ADP is released, and myosin is returned to its original state with no nucleotides bound (A). Cross-bridge cycling continues, with myosin "walking" toward the plus end of the actin filament, as long as Ca2+ is bound to troponin C.
Relaxationoccurs when Ca2+ is reaccumulated in the sarcoplasmic reticulum by the Ca2+ ATPase of the sarcoplasmic reticulum membrane) SERCA(
Rigor Mortis Several hours after death, all the muscles of the body go into a state ofcontracturecalled "rigor mortis"; that is, the muscles contract and become rigid, even without action potentials. This rigidity results from loss of all the ATP, which is required to cause separation of the cross-bridges from the actin filaments during the relaxation process. The muscles remain in rigor until the muscle proteins deteriorate about 15 to 25 hours later, which presumably results from autolysis caused by enzymes released from lysosomes. All these events occur more rapidly at higher temperatures
The Amount of Actin and Myosin Filament Overlap Determines Tension Developed by the Contracting Muscle
Effect of Muscle Length on Force of Contraction in the Whole Intact Muscle
Relation of Velocity of Contraction to Load when the load has been increased to equal the maximum force that the muscle can exert, the velocity of contraction becomes zero and no contraction results, despite activation of the muscle fiber.
Isometric Versus Isotonic Contraction Muscle contraction is said to beisometricwhen the muscle does not shorten during contraction andisotonicwhen it does shorten but the tension on the muscle remains constant throughout the contraction
Characteristics of Isometric Twitches Recorded from Different Muscles
Fast Versus Slow Muscle Fibers every muscle of the body is composed of a mixture of so-calledfastandslowmuscle fibers, with still other fibers gradated between these two extremes
Slow Fibers (Type 1, Red Muscle( • Smaller fibers. • innervated by smaller nerve fibers. • More extensive blood vessel system and capillaries to supply extra amounts of oxygen. • Greatly increased numbers of mitochondria, also to support high levels of oxidative metabolism. • Fibers contain large amounts of myoglobin, an iron-containing protein similar to hemoglobin in red blood cells. Myoglobin combines with oxygen and stores it until needed; this also greatly speeds oxygen transport to the mitochondria. The myoglobin gives the slow muscle a reddish appearance and the namered muscle.
Fast Fibers (Type II, White Muscle ) • Large fibers for great strength of contraction. • Extensive sarcoplasmic reticulum for rapid release of calcium ions to initiate contraction. • Large amounts of glycolytic enzymes for rapid release of energy by the glycolytic process. • Less extensive blood supply because oxidative metabolism is of secondary importance. • Fewer mitochondria, also because oxidative metabolism is secondary. A deficit of red myoglobin in fast muscle gives it the namewhite muscle.
Motor Unit- All the Muscle Fibers Innervated by a Single Nerve Fiber
Force Summation Summationmeans the adding together of individual twitch contractions to increase the intensity of overall muscle contraction. • Summation occurs in two ways: • by increasing the number of motor units contracting simultaneously, which is calledmultiple fiber summation,and • by increasing the frequency of contraction, which is calledfrequency summationand can lead totetanization.
1.Multiple Fiber Summation When the central nervous system sends a weak signal to contract a muscle, the smaller motor units of the muscle may be stimulated in preference to the larger motor units. Then, as the strength of the signal increases, larger and larger motor units begin to be excited as well, This is called thesize principle
Frequency Summation and Tetanization as the frequency increases, there comes a point where each new contraction occurs before the preceding one is over. As a result, the second contraction is added partially to the first, so the total strength of contraction rises progressively with increasing frequency. When the frequency reaches a critical level, the successive contractions eventually become so rapid that they fuse together and the whole muscle contraction appears to be completely smooth and continuous. This is calledtetanization.
MECHANISM OF TETANUS if the muscle is stimulated repeatedly, there is insufficient time for the sarcoplasmic reticulum to reaccumulate Ca2+, and the intracellular Ca2+ concentration never returns to the low levels that exist during relaxation. Instead, the level of intracellular Ca2+ concentration remains high, resulting in continued binding of Ca2+ to troponin C and continued cross-bridge cycling. In this state, there is a sustained contraction calledtetanus,rather than just a single twitch.
Skeletal Muscle Tone Even when muscles are at rest, a certain amount of tautness usually remains. This is calledmuscle tone. Because normal skeletal muscle fibers do not contract without an action potential to stimulate the fibers, skeletal muscle tone results entirely from a low rate of nerve impulses coming from the spinal cord
the same attractive forces between myosin and actin filaments cause contraction in smooth muscle as in skeletal muscle, but the internal physical arrangement of smooth muscle fibers is different.
Although most skeletal muscles contract and relax rapidly, most smooth muscle contraction is prolonged tonic contraction, sometimes lasting hours or even days. Comparison of Smooth Muscle Contraction and Skeletal Muscle Contraction • Slow Cycling of the Myosin Cross-Bridges • Low Energy Requirement to Sustain Smooth Muscle Contraction • Slowness of Onset of Contraction and Relaxation of the Total Smooth Muscle Tissue • Maximum Force of Contraction Is Often Greater in Smooth Muscle Than in Skeletal Muscle • "Latch" Mechanism Facilitates Prolonged Holding of Contractions of Smooth Muscle • Stress-Relaxation of Smooth Muscle
Regulation of Contraction by Calcium Ions smooth muscle does not contain troponin MYOSIN LIGHT CHAIN KINASE
smooth muscle can be stimulated to contract by multiple types of signals: by nervous signals, by hormonal stimulation, by stretch of the muscle, and in several other ways. • The principal reason for the difference is that the smooth muscle membrane contains many types of receptor proteins that can initiate the contractile process. • Still other receptor proteins inhibit smooth muscle contraction, which is another difference from skeletal muscle.
Probably half of all smooth muscle contraction is initiated by stimulatory factors acting directly on the smooth muscle contractile machinery and without action potentials. • Two types of non-nervous and nonaction potential stimulating factors often involved are • (1) local tissue chemical factors and • (2) various hormones
Source of Calcium Ions That Cause Contraction Through the Cell Membrane and from the Sarcoplasmic Reticulum Role of the Smooth Muscle Sarcoplasmic Reticulum Smooth Muscle Contraction Is Dependent on Extracellular Calcium Ion Concentration