Download

1 / 61
610 likes | 804 Views
The Molecular Basis of Inheritance. Campbell and Reece Chapter 16. The Search for Genetic Material. once Morgan proved genes are in chromosomes big debate started: Is the genetic material in chromosomes the DNA or the proteins? @ first case for proteins seemed stronger very heterogenous

E N D
The Molecular Basis of Inheritance Campbell and Reece Chapter 16
The Search for Genetic Material • once Morgan proved genes are in chromosomes big debate started: • Is the genetic material in chromosomes the DNA or the proteins? • @ first case for proteins seemed stronger • very heterogenous • great specificty
Evidence that DNA can Transform Bacteria 1928 Griffith studied Streptococcus pneumoniae
Transformation term coined by Griffith change in genotype & phenotype due to the assimilation of external DNA by a cell Avery spent the next 14 years identifying the “transforming agent”
Avery • & his colleagues announced DNA was the transforming agent • many were skeptical • Was bacterial DNA anything like eukaryotic DNA? • Nothing much known about DNA • Most scientists held to belief that proteins had to be transforming agent
Bacteriophages • are viruses that infect bacteria • “phages” for short • Virus made of a protein coat covering genetic material • to produce more viruses it must invade a cell & take over the cell’s metabolic machinery
Additional Evidence that DNA is Genetic Material • Chargraff • already knew DNA made up of: • Deoxyribose • Phosphate group • Nitrogenous Base (A, G, C, T) • analyzed DNA from # of species • 1950: base composition of DNA varies between species • made DNA more credible
Chargraff’s Rule • no matter what the source of DNA tested:
What is the structure of DNA? early 1950’s: scientists convinced DNA carried genetic info focus now on DNA’s structure knew arrangement of DNA’s covalent bonds
Watson & Crick Cambridge, England 2 young, unknown scientists same lab: Franklin & Wilkins doing x-ray crystallography on protein structure
X-Ray Crystallography of DNA Rosalind Franklin had purified some DNA and showed results to Watkins who was familiar with pattern made by a helical structure
Watson & Crick • began building models that satisfied: • known chemical properties of DNA • nitrogenous bases relatively hydrophobic • phosphate groups carry (-) charge • Chagraff’s rules • helical structure • how could this structure pass on genetic information?
DNA Structure antiparallel: arrangement of sugar-phosphate backbones in a DNA double helix means 1 strand runs 5’ 3’ going “up” * the other runs 5’ 3’ going “down”
DNA Structure because of size differences in dbl ringed purines vs. single ringed pyrimidines Watson & Crick knew could not have a purine linked with itself or the other purine also knew that adenine & thymine could form H bonds (2) with each other & cytosine & guanine could for 3 H bonds
Watson & Crick their 1 page paper published in Nature in April 1953 Watson, Crick, and Wilkins received Nobel Prize in 1962 (Franklin died in 1958)
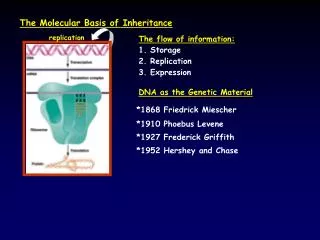
DNA Replication Watson and Crick’s 2nd paper stated their hypothesis on how DNA replicates: DNA model is pair of complimentary templates prior to replication H bonds broken & chains separate & unwind each chain then acts as template for formation onto itself of a new complimentary chain allows for exact duplication
Watson & Crick’s Semiconservative Model of DNA Replication predicts when a dbl helix replicates, each of the 2 daughter molecules will have 1 old strand and 1 new strand Conservative Model: 1 new daughter molecule with 2 new strands & the original molecule Dispersive Model: all 4 strands of DNA after replication have mixture of old & new parts
DNA Replication begins @ particular sites called: Origins of Replication short stretches of a specific sequence of nucleotides many bacterial loops of DNA have single origin proteins that initiate DNA replication recognize the sequence / attach to the DNA / separate the 2 strands by breaking H bonds creating “bubbles”
Replication Forks • @ each end of the replication bubble • Y-shaped region where DNA is unwinding • proteins that participate in the unwinding: • helicases • unwind double helix • single-strand binding proteins • bind to single strands prevents them from rewinding • topoisomerases • untwisting dbl helix puts strain on ahead of replication fork, these proteins relieve strain by breaking, swiveling, & rejoining DNA strands
Replication of DNA initial nucleotide chain made during DNA synthesis is actually a strand of RNA this RNA chain called a primer which is made by an enzyme called primase (last slide) primase starts a complementary RNA chain from a single RNA nucleotide then adds 1 @ time
Primers when primer 5 – 10 nucleotides long...new DNA strand will start from the 3’ end of the RNA primer
DNA Polymerase • enzyme that catalyzes the synthesis of new DNA by adding nucleotides to a pre-existing chain • 2 major one in prokaryotes • 11 different ones in eukaryotes • most require a primer & DNA template strand • rate: ~500 nucleotides/s in bacteria • ~ 50 nucleotides/s in human cells
Source of Nucleotides are in form of nucleoside triphosphates
Nucleoside Triphosphates are chemically reactive (like ATP, except sugar is deoxyribose, not ribose) as each nucleotide joins the growing end of a DNA strand 2 of the phosphate groups are lost as a molecule of inorganic phosphate in a couple exergonic reaction that drives the polymerization reaction
Antiparallel Elongation • each strand of DNA has directionality (1-way street) • & each strand oriented in opposite directions to each other • DNA polymerase III can add nucleotides only to the free 3’ end of a primer or growing DNA strand • along 1 template DNA polymerase synthesizes complementary strand continuously (5’ 3’ direction) • called the Leading Strand
Antiparallel Elongation • along opposite strand because of orientation, DNA polymerase III must work in direction away from the replication fork • called Lagging Strand • synthesized in short segments called: • Okazaki Fragments • ~ 1,000 – 2,000 nucleotides long in E. coli • ~ 100 – 200 nucleotides long in eukaryotes
DNA Replication Complex easy to think of DNA polymerase as a locomotive moving down template track but not really how it works: various proteins that participate in DNA replication form a large complex DNA replication complex doesn’t move, the DNA template moves thru the complex
Proofreading & Repairing DNA ~ 1/10 billion base pairs in completed DNA will be incorrect but right after strands replicated errors ~ 100,000 times more common DNA polymerases “proofread” each nucleotide against its template as soon as it is added when error found, incorrect nucleotide removed, correct 1 inserted
Proofreading & Repairing some errors evade DNA polymerase…other enzymes remove & replace incorrectly paired nucleotides some errors arise after replication: damage to DNA relatively common: usually corrected by b/4 becoming permanent mutations cells continuously monitor & repair damaged DNA
Repair Enzymes • ~100 in E. coli • ~ 130 in humans • most organisms use same mechanism to repair errors or damage • involves cutting out damaged area using DNA-cutting enzyme called nuclease • gap then filled with correct nucleotides done by a DNA polymerase & DNA ligase • 1 of these systems called nucleotide excision repair