Download

1 / 32
320 likes | 1.07k Views
The Visual System: From Eye to Cortex. Ch. 6. Outline. The Eye The Retina Visual Transduction by Rhodopsin From Retina to Primary Visual Cortex. The Eyes. Structure : review the major structures of the eye (diagram given in class)

E N D
Outline • The Eye • The Retina • Visual Transduction by Rhodopsin • From Retina to Primary Visual Cortex
The Eyes • Structure: review the major structures of the eye (diagram given in class) (note that the size of the pupils is a compromise between sensitivity and acuity)
The Eyes • ciliary muscles adjust the lenses to focus visual images sharply on each retina, regardless of the distance of the image from eyes; this focusing is called acommodation
The Eyes When we look at something near, the ciliary muscles contract, tension on the lenses is reduced, and they become more cylindrical • When we look at distant object, the muscles relax, tension on the lens is increased, and the lenses flatten
The Eyes • Binocular Disparity: unlike most vertebrates, most mammals have two eyes on the front of their heads, rather than one on each side; this cuts down the field of view, but it insures that most of what is seen is seen through both eyes
The Eyes • Because each eye sees things from a slightly different perspective, there is a difference in the two retinal images; this binocular disparity is greater for closer things; the degree of binocular disparity associated with a particular visual stimulus helps the brain create a 3-D perception from two 2-D retinal images thus depth perception is partially achieved from this (remember, we can still perceive depth with one eye closed)
The Retina • Structure: five layers receptor layer; horizontal cell layer; bipolar layer; amacrine cell layer; and retinal ganglion cell layer (diagram in class) • The receptor is the farthest from light; therefore, incoming light is distorted by four layers of neurons before reaching the receptors
The Retina • The fovea is a pit about 330 micrometers in diameter; it is the only part of the retina capable of mediating high-acuity vision. This is due, in part, to the fact that axons of retinal ganglion cells are thinnest over the fovea and light is distorted less before reaching the layer of receptors. The fovea is roughly in the center of your visual field.
The Retina • The optic disk is where the axons of retinal ganglion cells penetrate the retina and exit the eye; the optic disk has no receptors thus creating a blind spot
The Retina • We all have a blind spot at the optic disk, due to the exit of axons from the retinal ganglion cells
The Eyes • We are normally unaware of our blind spots, even when looking through one stationary eye because of completion; the visual system is able to use visual information gathered from receptors around the optic disk to complete the visual image • Completion illustrates the creativity of the visual system (demonstration in class)
The Retina • Cone and Rod Vision: in a sense, each of us has two visual systems; the photopic system functions in lighted conditions; the scoptopic system functions in dim light
The Retina • Cones mediate photopic vision; rods mediate scotopic vision; this is called the duplexity theory of vision • The photopic system mediates high-acuity vision; the scotopic system mediates low-acuity vision
The Retina • The photopic system has low sensitivity with few receptors’ information combined at the next cell level (low convergence) • The scotopic system has high sensitivity with many receptors converging on ganglion cells (high convergence) • Only cones in the fovea; rods predominate in the periphery
The Retina • Our eyes are in continual motion; they make a series of fixations (about 3 per second) connected by saccades (rapid movements between fixations)
The Retina • Eye movements keep the visual image in continual motion on the retina; the importance of this movement is illustrated by the fact that stabilized retinal images disappear; most visual system neurons respond to change, not to steady input
The Retina • What we perceive at any instance is the sum of the input that has been received during the last few fixations; this temporal integration of the retinal images explains why our visual images are detailed, colored, and wide-angled, despite the small size of the fovea; it also explains why things don’t disappear when we blink
Visual Transductionby Rhodopsin • Tranduction is the conversion of one form of energy to another; the first step in visual transduction is conversion of light to neural signals by the rods and cones
Visual Transductionby Rhodopsin • Transduction by rods is well understood; there is pink substance in rods called rhodopsin; light is absorbed by rhodopsin, and this bleaches it (removes its color); this bleaching occurs because retinal and opsin, the two components of rhodopsin, separate in light
Visual Transductionby Rhodopsin • This bleaching reaction hyperpolarizes the rods, thus the bleaching reaction tranduces light to a cascade of intracellular events resulting in neural signal • When rhodopsin is totally bleached by intense light, rods lose their ability to absorb light and transduce; however, in the dark, retinal and opsin are reunited, and rhodopsin regains its color and its ability to absorb light and transduce
Visual Transductionby Rhodopsin • Rhodopsin is a G-protein linked receptor that responds to light by initiating a cascade of intracellular chemical events • cGMP is an intracellular chemical that keeps Na+ channels partly open when rods are in darkness; rods are thus slightly depolarized
Visual Transductionby Rhodopsin • Bleaching rods via exposure to light results in an intracellular cascade of events that deactivates cGMP, closing Na+ channels, which hyperpolarizes the rod and reduces the release of glutamate • Rods thus transmit signals through the neural system via inhibition
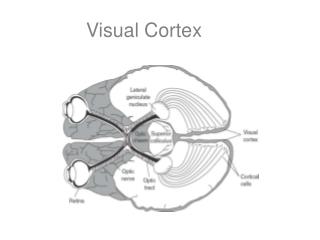
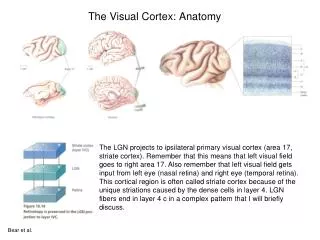
From Retina toPrimary Visual Cortex • This is called the retina-geniculate-striate pathway (diagram in class) • The primary visual cortex is striate cortex after the stripe seen in a Nissl-stained cross section of lower layer IV; the stripe is composed of the terminals of axons from the lateral geniculate nuclei
From Retina toPrimary Visual Cortex • The retinal ganglion cell axons from the nasal hemiretinas decussate via the optic chiasm (they are contralateral); those from the temporal hemiretinas remain ipsilateral providing redundancy for the system
From Retina toPrimary Visual Cortex • The most important feature of visual cortex organization is its retinotopic layout; the surface of the visual cortex is a map of the retina
From Retina toPrimary Visual Cortex • The retina-geniculate-striate system includes two largely independent channels: The P pathway and the M pathway
From Retina toPrimary Visual Cortex • Parvocellular layers (P pathway) are found in the top four layers of each lateral geniculate nucleus (LGN) and are composed of small body neurons; they are responsive to color, fine detail patterns, and react to slow or stationary objects • Magnocellular layers (M pathway) are found in the bottom two layers of the LGN, composed of large body neurons, and are responsive to rods and movements