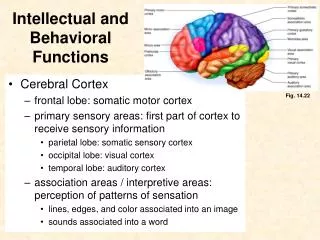
Download
1 / 13
250 likes | 687 Views
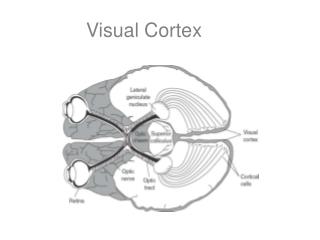
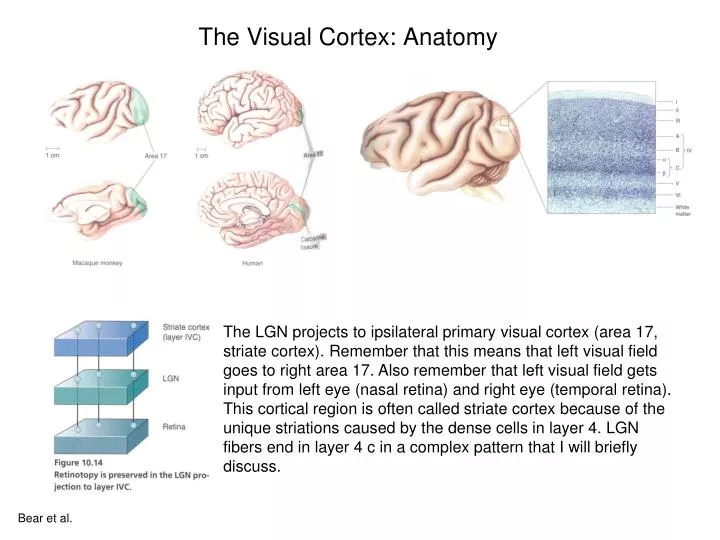
The Visual Cortex: Anatomy. The LGN projects to ipsilateral primary visual cortex (area 17, striate cortex). Remember that this means that left visual field goes to right area 17. Also remember that left visual field gets input from left eye (nasal retina) and right eye (temporal retina).

E N D
The Visual Cortex: Anatomy The LGN projects to ipsilateral primary visual cortex (area 17, striate cortex). Remember that this means that left visual field goes to right area 17. Also remember that left visual field gets input from left eye (nasal retina) and right eye (temporal retina). This cortical region is often called striate cortex because of the unique striations caused by the dense cells in layer 4. LGN fibers end in layer 4 c in a complex pattern that I will briefly discuss. Bear et al.
Bear et al. The Visual Cortex: Circuitry Pyramidal cells (excitatory) of area 17. The stellate cells of layer 4 C are unique modified pyramidal cells. It is important to remember that there are many classes of inhibitory (GABA) interneurons that are not shown here. There are perhaps as many as 30 interneurons in cortex. Upper cortical layers project to cortex; in this case to areas 18,19. Lower layers (5,6) project to brainstem and thalamus. Layer 5 projects to superior colliculus (control of eye movements). Layer 6 projects topographically to LGN. LGN projects to layer 4 stellate cells. These cells project to cells in the upper layer; cells in the upper layers project to the lower layers and out. Note the vertical (columnar) organization. This is a highly simplified view of cortical circuitry.
Bear et al. The Visual Cortex: Spatial Organization 1 The above represents the projection of one eye to the striate cortex (layer 4C). Note the stripes. The blank regions are the stripes representing the other eye. So, in layer 4C, the eyes representing a visual field remain separated: because this separation extends through a vertical column, these are sometimes called the oculur dominance columns. A stain for cytochrome oxidase reveals an unexpected columnar organization: the CO blobs. It is independent of the OD columns. We’ll see the blobs functional significance later on.
The Visual Cortex: Spatial Organization 2 Bear et al The M (movement) and P (high resolution) pathways are kept separate in striate cortex. Both pathways converge on the blobs. Note the vertical organization of the intrinsic circuitry. Now what does the cortex do with this input?
The Visual Cortex: Functional Organization- Simple Cells Many pyramidal neurons in striate cortex respond to oriented moving bars and not to spots of light (remember that LGN relay cells respond to spots of light. So there are two important transformations that occur in striate cortex: Most cells are selective for a particular orientation and they require some movement. Orientation selectivity depends on the convergence of input from an appropriate subset of LGN relay cells as indicated above. Orientation selectivity may also be enhanced by cortical circuitry. The cell illustrated here responds to an oriented bar in one region of space; this type of cell is called a simple cell. Bear et al.
The Visual Cortex: Functional Organization- Complex Cells Bear et al. Complex cells are typically found in the upper and lower layers of striate cortex. They are very varied in their properties but always respond to oriented moving bars. The difference between complex cells and simple cells is that complex cells respond over a wide area. Some complex cells also respond to to movement in one direction only.
The Visual Cortex: Overall Organization Bear et al. The figure to the left shows optical imaging of ocular dominance columns (mid) and orientation columns (bottom). You can see that the orientation stripes cut through the OD stripes. The figure at the top shows the conceptual diagram of this relationship. For each bit of striate cortex (a small region of the visual field) each eye representation has complete representation of all possible orientations (layer 4C); the upper layers combine these representions to allow the visual system to estimate depth (stereopsis). The blobs represent a completely different information channel. Cells in the blobs are not sensitive to orientation or movement. The blobs neurons respond to colour (primates); this channel is not found in cats, rodents etc.
Plasticity in the Visual Cortex 1 Ocular dominance stripes with normal rearing. Ocular dominance stripes with one eye occluded during a critical period. The occluded eye does not capture as much cortical territory as the normal eye. This molecular biology of this effect has been intensely studied. A variety of factors, including neurotrophins, are involved but a complete picture is lacking. Kandel et al.
Plasticity in the Visual Cortex 2 Kandel et al. This developmental sequence can be disrupted by eye closure and depends on complex molecular interactions driven by correlated input from the two eyes. Development of ocular dominance columns in cats.
Plasticity in the Visual System: Competition for Targets Normal amphibian tectum: tracer injection in the contralateral eye. Amphibian tectum with third eye grafted on: the two eyes compete for tectal target space and produce a columnar arrangement- artificial ocular dominance. NMDA receptors are critically involved in this process. Kandel et al.
The Visual Cortex: Estimating Depth Kandel et al. Objects at the eyes fixation point are fused; those behind or in front of the fixation point (retinal disparity) are interpreted as closer or further away from you. A population of cells in striate cortex combines the images from the 2 eyes in such a way that they respond to retinal disparity and thus signal the relative depth of objects. This system is very important for many animals such as carnivores (estimating distance to prey) and primates (estimating distance to insect or fruit).
The Visual Cortex: Projections- Two Processing Streams 1 Bear et al. There are two main processing streams for primate vision. A dorsal stream goes through the middle temporal lobe and onto the parietal cortex. It deals with movement (the M channel to a large extent) and is used for navigation and motion perception. The ventral stream goes into the temporal lobe and deals with identification of visual input (the P channel is a big contributor).
The Visual Cortex: Projections- Two Processing Streams 2 Bear et al. Kandel et al. Response of a neuron in temporal cortex (monkey, ventral stream) to faces. Recently far more specific cells have been found in humans- responding to pictures of Bill Clinton etc. Specific cortical areas respond to such “communication” signals. PET image of activation of the dorsal stream in response to visual motion. Different neurons in the visual system respond to different features: motion, shape, colour etc. How are these different attributes combined to form a single percept- this is the binding problem. One controversial hypothesis is that synchronized activity in neurons representing these different features are the basis of the unity of perception.