Download

1 / 19
190 likes | 359 Views
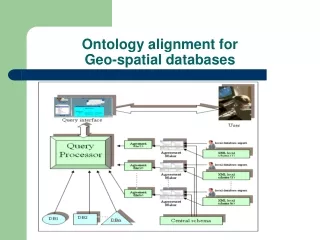
Initial Proposal for the RNA Alignment Ontology. Rob Knight Dept Chem & Biochem CU Boulder. What do we want to do?. Represent detailed structural info and other metadata on alignment Avoid horizontal and vertical expansion Explicitly annotate correspondences at the level where they occur.

E N D
Initial Proposal for the RNA Alignment Ontology Rob Knight Dept Chem & Biochem CU Boulder
What do we want to do? • Represent detailed structural info and other metadata on alignment • Avoid horizontal and vertical expansion • Explicitly annotate correspondences at the level where they occur
…so real alignments look like this, to shoehorn everything into columns that are assumed to be homologous
Homology is problematic… • Fundamental problem: systems that are homologous at one level are not necessarily homologous at other levels • E.g. bat wings and bird wings: homologous as pentadactyl limbs, but not homologous as wings • Homology is hierarchical andcan partially overlap at any level(e.g. Griffiths 2006) Frog forelimbs Bird forelimbs Rodent forelimbs Bat forelimbs Mammal forelimbs Tetrapod forelimbs Ridley “Evolution” 3rd ed.
…and correspondence need not be homology at all! • Example from SELEX: hammerhead ribozymes independently evolved at least three times: in nature, and in Jack Szostak and Ron Breaker’s labs • However, we still want to be able to align the functionally equivalent sequences although there is not evolutionary relationship
Problem: have millions of fragments, want to align (incl. noncanonical pairs) + assign named regions
Solution • Use existing alignment, try to fit new seqs in • Would be improved if we could explicitly annotate helices, noncanonical pairs, etc. on the sequence overall • For display, need to easily show/hide groups of sequences and/or regions of the sequence
Use case 2: SELEX • From large number of unaligned sequences, want to identify motifs like this (Majerfeld & Yarus 2005)
How is this currently done? • Find regions that are similar in more sequences than chance • Group these sequences centered on the “motif” • See if the parts of the motif can be related by helices • See if anything else is reliably found by the motif • Repeat for other families and see if there are relationships between them • Group these families together, then iterate
So how do we handle all this? A proposal Entities: • sequence_region: a thing that defines a set of bases relative to some sequence (i.e. with indices for each base) • paired_sequence_region: two regions linked by pairs • helical_sequence_region: two regions completely paired • base: region that consists of single nucleotide • base_pair: region that consists of two, paired bases • canonical_base_pair: base pair that is cis-WW • loop: contiguous sequence_region stretching from i to j such that i-1 and j+1 are a base pair • etc. (bulge, internal_loop, junction, etc.)
So how do we handle all this? A proposal Relationships: • correspondence: relation among set of sequence_regions implying all share a feature (with metadata about how determined) • homology: correspondence implying continuous chain of descent preserving the relation • sequence_similarity: correspondence implying regions are similar in primary sequence • two_d_structure_similarity: correspondence implying regions are similar in 2D structure, i.e. nested canonical base pairs • secondary_structure_similarity: correspondence implying regions are similar in secondary structure, i.e. incl. pseudoknots/noncanonicals • tertiary_structure_similarity: correspondence implying regions are similar in 3D structure
So how do we handle all this? A proposal Relationships: • pairing: relation that asserts that two sequence_regions each have parts of at least one base_pair that connects them • helical_pairing: pairing that includes several base_pairs (not necessarily contiguous) between two sequence_regions • unbroken_helical_pairing: helical_pairing that includes no bases in the sequence_regions that are not paired with the other sequence_region, in order • base_pairing: pairing that connects exactly two bases, annotated with the Leontis-Westhof classification More exotic uses for alignment: • microrna_target: pairing relation in which one member is a miRNA and the other is an mRNA according to SO • same_microrna_target: a relation among a set of sequences that have microrna_target relation to the same miRNA
Implementation notes • Must be able to name regions (e.g. P3 in RNaseP) and subclass them (e.g. P3 in firmicutes) • Must be able to subclass homologies, e.g. homologous as wing vs. homologous as limb • Correspondences are all symmetric and transitive, so can implement as set of regions that share the correspondence • (probably) don’t want to reify names of parts of well-known RNAs in the overall RNAO?
Acknowledgements Knight Lab members: • Cathy Lozupone • Micah Hamady • Chris Lauber • Jesse Zaneveld • Jeremy Widmann • Elizabeth Costello • Jens Reeder • Daniel McDonald • Anh Vu • Ryan Kennedy • Julia Goodrich • Meg Pirrung • Reece Gesumaria Trp project: • Irene Majerfeld • Jana Chochosolousova • Vikas Malaiya • Matthew Iyer • Mike Yarus RNA Alignment Ontology working group: • James. W. Brown • Fabrice Jossinet • Rym Kachouri • B. Franz. Lang • Neocles Lenotis • Gerhard Steger • Jesse Stombaugh • Eric Westhof Other coauthors: • Amanda Birmingham • Paul Griffiths • Franz Lang NSF RCN grant # 0443508