Download
1 / 1
10 likes | 125 Views
Effect of time-delay on reaction-diffusion wavefronts modeling biological invasions Daniel Campos 1 , Joaquim Fort 2 , Vicenç Méndez 3 and Vicente Ortega-Cejas 1

E N D
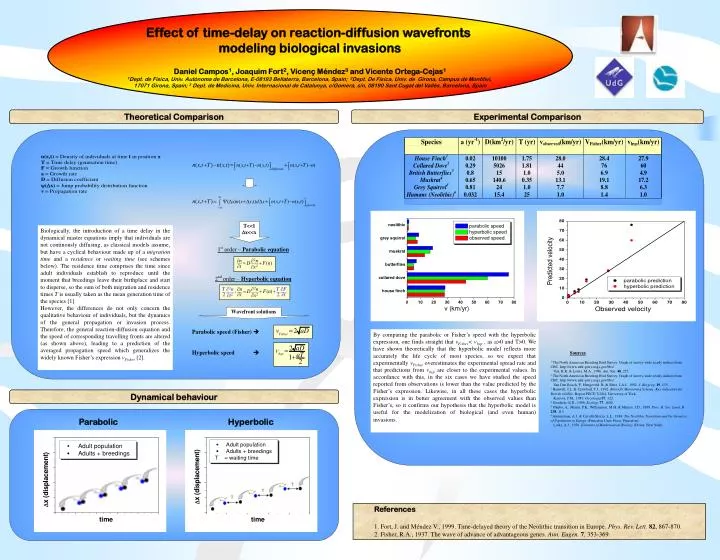
Effect of time-delay on reaction-diffusion wavefronts modeling biological invasions Daniel Campos1, Joaquim Fort2, Vicenç Méndez3 and Vicente Ortega-Cejas1 1Dept. de Física, Univ. Autònoma de Barcelona, E-08193 Bellaterra, Barcelona, Spain; 2Dept. De Física, Univ. de Girona, Campus de Montilivi, 17071 Girona, Spain; 3 Dept. de Medicina, Univ. Internacional de Catalunya, c/Gomera, s/n, 08190 Sant Cugat del Vallés, Barcelona, Spain Theoretical Comparison Experimental Comparison Biologically, the introduction of a time delay in the dynamical master equations imply that individuals are not continously diffusing, as classical models assume, but have a cyclical behaviour made up of a migration time and a residence or waiting time (see schemes below). The residence time comprises the time since adult individuals establish to reproduce until the moment that breedings leave their birthplace and start to disperse, so the sum of both migration and residence times T is usually taken as the mean generation time of the species [1]. However, the differences do not only concern the qualitative behaviour of individuals, but the dynamics of the general propagation or invasion process. Therefore, the general reaction-diffusion equation and the speed of corresponding travelling fronts are altered (as shown above), leading to a prediction of the averaged propagation speed which generalizes the widely known Fisher’s expression vFisher [2]. By comparing the parabolic or Fisher’s speed with the hyperbolic expression, one finds straight that vFisher<vhyp , as a>0 and T>0. We have shown theoretically that the hyperbolic model reflects more accurately the life cycle of most species, so we expect that experimentally vFisher overestimates the experimental spread rate and that predictions from vhyp are closer to the experimental values. In accordance with this, in the six cases we have studied the speed reported from observations is lower than the value predicted by the Fisher’s expression. Likewise, in all these cases the hyperbolic expression is in better agreement with the observed values than Fisher’s, so it confirms our hypothesis that the hyperbolic model is useful for the modelization of biological (and even human) invasions. • Sources • 1 The North American Breeding Bird Survey. Graph of survey-wide yearly indices from CBC. http://www.mbr-pwrc.usgs.gov/bbs/. • Veit, R.R. & Lewis, M.A., 1996. Am. Nat.48, 255. • 2 The North American Breeding Bird Survey. Graph of survey-wide yearly indices from CBC. http://www.mbr-pwrc.usgs.gov/bbs/. • Van Den Bosch, F., Hengeveld, R. & Metz, J.A.J., 1992. J. Biogeog.19, 135. • 3 Banwell, J.L. & Crawford, T.J., 1992. Butterfly Monitoring Scheme. Key indicators for British wildlife, Report PECD 7/2/84, University of York. • Kareiva, P.M., 1983. Oecologia57, 322. • 4 Grosholz, E.D., 1996. Ecology77, 1680. • 5 Okubo, A., Miami, P.K., Williamson, M.H. & Murray, J.D., 1989. Proc. R. Soc. Lond. B238, 113 • 6 Ammerman, A.J. & Cavalli-Sforza, L.L., 1984. The Neolithic Transition and the Genetics of Population in Europe (Princeton Univ. Press, Princeton). • Lotka, A.J., 1956. Elements of Mathematical Biology (Dover, New York). Dynamical behaviour Parabolic Hyperbolic References 1. Fort, J. and Méndez V., 1999. Time-delayed theory of the Neolithic transition in Europe. Phys. Rev. Lett.82, 867-870. 2. Fisher, R.A., 1937. The wave of advance of advantageous genes. Ann. Eugen.7, 353-369.