Download

1 / 29
290 likes | 421 Views
Viruses. Some Random Statistics 1918-1919 Spanish Flu (A/H1N1) 20-40 million deaths world-wide 1957 A sian (A/H2N2) & 1968 Hong-Kong (A/H3N2) 1.5 million+ deaths ~ 35 – 50 million infected with influenza (USA) each year ~ 20,000 deaths/year

E N D
Some Random Statistics 1918-1919 Spanish Flu (A/H1N1) 20-40 million deaths world-wide 1957 Asian (A/H2N2) & 1968 Hong-Kong (A/H3N2) 1.5 million+ deaths ~ 35 – 50 million infected with influenza (USA) each year ~ 20,000 deaths/year ~ 64,000 deaths/year due to influenza + pneumonia Hantavirus (flu-like symptoms which may progress to severe difficulty in breathing & death) ~ 50% mortality (USA- Four corners, Canadian prairies) HIV ~ 30 million infected worldwide ~ 100% mortality Ebola (fever, headache, joint-muscle aches, weakness progressing to diarrhea, vomiting, internal/external bleeding) ~ 90% mortality West Nile (USA) (flu-like symptoms) 2002: 3389 infected 201 deaths... 5.9% 2003: 9862 infected 264 deaths... 2.6 2004: 2539 infected 100 deaths... 3.9 2005: 3000 infected 119 deaths... 3.9 2006: 4269 infected 177 deaths... 4.4 2007: 3623 infected 124 deaths... 3.4
Measles ~ 30+ million cases world-wide with ~ 1 million deaths Hepatitis A ~ 1.4 million infections world-wide Hepatitis B (chronic liver inflammation, cirrhosis, cancer) ~ 2 BILLION people infected (total) world-wide ~ 10% develop liver cancer Hepatitis C (chronic liver inflammation, cirrhosis, cancer) ~ 3 – 4 million infected world-wide ~ 10% develop liver cancer Rotavirus (vomiting & severe diarrhea) ~ 600,000 children die Cervical Cancer ~50% due to HPV ~ 470,000 million new cases world-wide ~ 230,000 deaths Obviously viruses are an important contributor to human disease and mortality
There are more varieties of viruses than any other biological entity on the planet; simply millions of different ones. Most, however, are irrelevant to human disease because most are incapable of infecting humans. Those that do, however, are capable of causing a devastating array of diseases; from the common cold and influenza to Ebola, hepatitis, and AIDS. Like any other pathogen, they illicit an immune response that then eliminates the virus; providing, of course, that the virus is recognized as foreign, and that the genes exist that “allow” us to develop immunity. As inferred, an inability to develop immunity to the small pox virus probably wiped out as many as 80% of the native population in North America, allowing the Europeans to dominate and subjugate the remaining population. AIDS, on the other hand, develops because it is only the exceedingly rare human that is capable of recognizing the fast-mutating HIV and amazingly evasive HIV virus. As recent research indicates, it is possible to develop immunity against the virus from a carefully crafted vaccine; indicating that humans possibly can develop immunity against HIV but it is the fast mutation rate and “evasive” abilities of the virus that “prevents” immune recognition. As the list on the previous slides attests, a lot of people die as a result of viral infections. And from the West Nile data... It appears that lots of publicity and education about avoiding mosquitoes and early diagnosis can reduce infections and deaths...
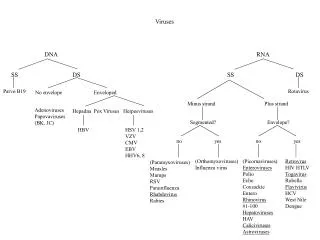
Types of Viruses DNA Viruses ds-DNA ss-DNA Smallpox Parovirus B19 Herpes Adenovirus Human Papillomavirus RNA Viruses antisense RNA sense RNA Measles Polio Mumps Rhinovirus Ebola Corona Virus Influenza West Nile Virus Rabies Ruebella diploid ssRNA Retrovirus HIV HTLV
There are different variations of viruses based on the form of their genetic material. DNA viruses such as the smallpox, herpes, and human papillomavirus have their genetic material in the form of double-stranded DNA; just like ours. When the DNA is injected into the cell it is picked up by the normal enzymes for transcription and transcribed into mRNA and then translated into viral proteins by the normal protein synthesis machinery that is already active in the cell. RNA viruses such as measles, mumps, ebola, influenza and rabies have their genetic material in the form of antisense RNA while the rhinovirus, corona virus, ruebella virus, and West Nile virus have their genetic material in the form of sense RNA. RNA retroviruses have genetic material in the form of diploid, single-stranded RNA that must first be reverse-transcribed into double-stranded DNA (DNA sequence converted into RNA sequence is transcription so RNA sequence to DNA sequence must be reverse transcription) which is then handled like any other DNA-based genes. The resulting DNA from most retroviruses is integrated into the DNA of the host. The enzyme to do this is supplied by the virus; hence the name retrovirus: meaning reverse transcriptase virus (or something like that). The human immunodeficiency virus is an example of a retro virus. This virus infects the helper T-cells and ultimately can greatly impair their function and reduce their numbers to ineffective amounts. This would explain why HIV infections can lead to autoimmune deficiency disease; an individual can’t develop immunity toward any new infection and very often dies due to complications from infectious diseases that normally would not be fatal. Most viruses are pretty selective in terms of their ability to infect specific types of cell. For example, there are viruses that can only infect bacteria cells (bacteriophages) and some viruses that can only infect upper respiratory epithelial cells (influenza virus, rhino virus), or sensory nerve cells (varicella), and so on. The virus capsid has receptor-type proteins on the outer surface that can recognize only one type of cell on the basis of the membrane-bound proteins or phospholipids which exist on the cell surface. For example, the HA protein of the influenza virus recognizes the sialic acid component of a sialyl-lactose glycolipid (a membrane phospholipid). If the virus mutates and starts producing a capsid protein that recognizes a different cell type, then the virus will be able to invade a new cell type. This has happened with a strain of avian (bird) influenza virus in Hong Kong in 1997; the HA protein of a avian influenza virus mutated to recognize the sialyl-lactose configuration of human cells and caused an epidemic of influenza. Sometimes simply being exposed to mass quantities of a particular non-human virus can still lead to infection. An example of this would be the A/H5N1 strain of avian influenza virus
Viruses do not have the capacity to reproduce because they do not possess the genes necessary to code for all of the appropriate proteins. They have a minimal genome sufficient only for replicating capsid proteins, viral genome, and viral-specific enzymes. They must infect other cells and use the cellular array of protein-synthesis enzymes to reproduce the viral proteins and genome and assemble new viruses.
Viruses are strange creatures which are barely alive. They do not fit the normal definition of living because they cannot reproduce on their own. They need another living species to do that for them. A virus consists of a protein capsule called a capsid, inside of which are contained the viral genes in the form of DNA, or RNA as well as possibly an enzyme or two (or more) depending of which virus we are talking about. The only function of a virus is to bind to a living cell of some sort (specific viruses can bind only to specific cell types with the proper receptor or receptors) and inject its contents into the cell. Once the viral contents are inside the cell then the viral genes are translated into the viral proteins by the normal protein synthesis machinery that already exists inside the cell. When all of the viral proteins are synthesized they are assembled into new viruses. Many new copies of the virus will be made by the cell and the newly made viruses then are released from the cell or they break out of the cell and then diffuse around to infect other nearby cells with the same surface receptors. Obviously because some the viruses break out of the cell, the cell membrane will be ruptured. Either the cell dies and initiates an acute inflammatory response (or enough cells die that the organ fails and the infected person dies) or the damage is not severe enough to kill the cell but an inflammatory response in initiated anyway. Very often it is the inflammatory response to the virus-induced damage that produces the symptoms of the infection. Any resulting cell death from the infection can possibly lead to more serious consequences if the damage is extensive enough (organ dysfunction or failure). In some instances the viruses exit through a budding mechanism which does not kill or even damage the cell. Some of these viruses even get encapsulated with the cell membrane and therefore acquire the ability to evade immune surveillance (human cell membrane coat = no immune response).
Viruses attach to specific cells via their docking proteins. For example: Influenza - HA: recognizes sialic acid on glycoproteins /glycolipids Rhinovirus: ICAM-1 receptor/LDL receptor HIV: CD4 receptor of hT-cells/macrophages Measles: CD46 receptor (inhibits complement activation) & SLAM (Signaling Lymphocyte Activation Molecule) of T- cells In many cases, once inside, the viral contents are released into the cell and the genome is incorporated into the nucleus and the cells protein synthesis machinery is “hijacked” to replicate the viral proteins. Viruses supply their own specialized enzymes to ensure that the replication process proceeds if the cell cannot handle the specific form of viral genome.
After the viruses have been replicated by the cell, the viral particles either rupture the cell membrane to break out of the cell or fuse with the membrane and break off as a cell-membrane-enveloped particle. These processes will either kill the cell or injure the cell. In many cases, a local inflammatory response (please see PPt: HumanDisease-2-ImmuneFunction) is initiated as a result of the viral release. In some cases viral release doesn’t injure the cells but, rather, induces release of inflammatory-related cytokines
A little more on avian influenza: the avian influenza viruses of type A are classified by the particular type of haemagglutinin protein (15 different subtypes) and neuraminidase protein (9 different subtypes) on their surface. The influenza A/H5N1 strain of avian influenza is a particularly virulent strain of virus which is highly infective in birds and the only one which causes severe/fatal disease in humans. Because many humans happen to work with large numbers of birds (poultry farmers for example) it is possible that some humans may be exposed to huge numbers of this strain if they closely handle infected birds. This, of course has happened over the last 10 years where by late fall of the year 2005 about 100 people who have had very close contact with large numbers of infected poultry have become infected with the A/H5N1 virus. Unfortunately, this strain of virus is highly pathogenic and there is about a 25% to 80% fatality rate (depending on where the outbreak occurred) if infected. Why the avain H5N1 doesn’t easily infect humans is because the H5N1 subtype prefers a sialic acid joined to lactose with an α 2-3 linkage which is observed only in ciliated respiratory cells of humans (think mucus here) but in a very high percentage of intestinal cells of birds. Thus it takes exposure to large numbers of A/H5N1 virus to produce an effective infection in humans. On the other hand, if the HA protein of the A/H5N1 virus mutates to recognize the much more common sialic acid version (α 2-6 linkage) of human respiratory cells than it will become very easy to be infected by this virus. This is, of course a major fear today and governments all over the world are finally trying to do something to protect their citizens from possible infection by a highly virulent virus which has not been seen in humans before (no one has any immunity so it can be both deadly and spread very fast).
Human flu, on the other hand is commonly caused by a similar Type A influenza virus but there are only 3 known H proteins and 2 known N proteins. The seasonal form of influenza is typically the A/H3N2 or the A/H1N1 (and the Type B virus), where small changes in the genes of these viruses resulting from genetic drift produce slight changes in the H3 (or H1/H2 or N2/N1) proteins, producing a strain (same strain) of virus that is no longer recognized (or recognized not very well, ie. some cross-reactivity) by antibodies produced from the previous infection by the same (now, slightly different) strain. Type A/H1N1 strains have been responsible for the massive influenza pandemic of 1918-1919 where tens of millions of people died worldwide: 2.7% fatality rate compared to 0.1% for seasonal flu and about 0.5% to 1% for the 2009 A/H1N1 strain. The new A/H1N1 strain is an interesting virus because it has a unique genetic heritage. The RNA genes of this virus include genes from a North American avian H1N1, a North American version of swine H1N1, a human A/H1N1 and a couple non-North American swine A/H1N1 viruses. This combination comes about from a process called genetic shift. For this to happen, an animal (or human) needs to be infected with 2 different influenza viruses at the same time. When the 2 viruses “inject” their RNA into the host cells at the same time, then there can be a re-combination of the viral genes to produce H1N1 proteins that are very much different from the “parent” proteins. Apparently, this genetic shift has happened several times to result in the current A/H1N1 strain, a strain that most people have partial immunity against since the last H1N1 pandemic was in the ‘50s.
Specific symptoms of a viral infection depend on the specific type of cells infected & on the severity of cellular damage or extent of altered cell function. An example of differing symptoms from different viruses can be illustrated with the rhinovirus and influenza virus. Both are families of viruses that are highly infective in humans and are not (usually) fatal. Both infect the upper respiratory epithelia but lead to a different (but overlapping) array of symptoms due to the different degrees of damage and inflammation in response to the virus infections and to the different (but overlapping) array of cells infected. The HA (haemagglutinin) surface protein of influenza viruses bind to sialic acid residues of membrane lipoprotein polysaccharides while the rhino virus binds to the ICAM receptor (or LDL receptor). One major difference in ability to grow is that the rhinovirus likes temperatures ~33°C while the influenza virus likes temperatures a little warmer (34-35°C). This is probably one reason why rhinoviruses infect the nose and sinuses (they are cooler) much more while influenza virus infects the warmer lungs much more. A cold rarely causes a fever or headache while the “flu” always does; a cold rarely causes aches and pains and weakness and never causes exhaustion while the flu causes severe exhaustion, weakness, and lots of aches and pains. A major reason for these different symptoms is that the influenza virus causes extensive damage to the infected cells when they break out of them. The influenza virus also stimulates the infected cells to release interferon. The interferon is the inflammatory cytokine which causes fever and is largely responsible for the fatigue and aches and pains which accompany an influenza infection. On the other hand, the rhinovirus does not cause cellular damage but only induces the infected cells to release histamine. The stuffy and runny nose and sinus congestion are a result of the local effects of histamine release from the infected cells.
Herpes Virus HHV-1 (Herpes Simplex Virus I) virus infects oral skin epithelial/mucosal cells, multiplies within them & breaks out, causing blisters; some viral particles travel through sensory nerves to ganglion where they become inert. Later activation results in secondary lesions… HHV-2 (Herpes Simplex Virus II) same as above except more infectious in genital epithelia/mucosa. HHV-3 (Varicella zoster Virus) same as above except more infectious to skin epithelia on upper body resulting in chickenpox. Secondary activation results in shingles. HHV-4 (Epstein-Barr Virus) associated with Burkett Lymphoma, nasopharyngeal cancer, and mononucleosis. HHV-8 (Human Herpesvirus 8 - Kaposi’s sarcoma-associated) Herpes type viruses comprise one of the more common families of viruses which infect humans. Herpes simplex I, II, or III probably infect 80% (and most likely everyone) of the world. They cause cold-sore like skin eruptions and the Herpes Simplex I is mostly responsible for cold-sores on the face and lips, Simplex II is mostly responsible for genital herpes, and Simplex III (Varicella Zoster) is mostly responsible for the Chicken Pox, and if the virus is re-activated, for Shingles. There is really no cure for any of the Herpes infections although there is a vaccine available for Varicella Zoster (since 1995). The virus infects skin epithelial cells and on the first “outbreak”, the viral particles infect sensory nerves in the different areas (I = mostly face, II = mostly genitals, III = mostly skin of trunk and scalp) and lay dormant in the ganglions of the infected nerves. Many years later an outbreak can occur again, resulting in a re-appearance of the cold-sore like lesions (I & II) or in the severe pain and rash of Shingles (III). Epstein-Barr virus (EBV) (also one of the Herpes viruses) is responsible for infectious mononucleosis. As with the other Herpes viruses there is no known prevention (except for the varicella zoster one, of course) other than avoiding contact with infected persons and no treatment other than treating the symptoms. There are no drugs which can directly kill viruses!
Human Papilloma Virus HPV provides a nice example of how a virus can produce different symptoms based on the cell type infected and the specific cellular function which is altered. There are over 80 different types of this DNA virus; some of which cause warts, and in some cases, cervical cancer. To see how this works a little information on cell division and control of cell division is necessary. The normal progression from a single cell through the cell-division process is called the Cell Cycle. Human papilloma virus (HPV) is actually a family of viruses (more than 80 different strains) that cause benign tumors of the epidermis; commonly called warts. Some strains of the HPV cause cancer; for example cervical cancer is caused by this little beast. How HPV does this is a rather interesting story. In order to cause a tumor, the virus must somehow increase rates of cell division. The epithelial cells of the epidermal tissue (whether it be skin, oral mucosa, or genital mucosa) are infected by the virus which then is replicated by the infected cells. Normally this would end up causing some sort of damage to the cells membrane when the viral particles leave. HPV is a weird virus which requires cells to divide in order to replicate the virus. The virus does this in a very clever way.
Cell Division Signals Cancer cells continuously divide in an uncontrolled fashion DNA Damage “Promotion” “Initiation” Normal cells enter the cell cycle only when stimulated by “well-controlled” growth signals. “Progression”
Normally cells will not divide. They are in what is called G0 Arrest (Growth = Zero, arrest). Most cells also are terminally differentiated. This means that they are no longer immature, small, all-cells-look-the-same-and-can-turn-into-any-other cell-stem cells, but rather, grown up and look like mature tissue cells (and different from other tissue cells). A normal cell-division cycle would be G0 (the cell is quiescent) - progresses into G1 (grows to be a bigger cell) - which progresses into S Phase (all of the DNA is duplicated so the cell can divide into two cells; each with its own complete set of DNA) - which progresses into the G2 phase (grows again so there is enough of the larger cell to make two complete new cells when it divides) - which finally progresses into the M phase (mitosis, or division). If the original cell was a stem cell then one of the two new cells differentiates into a tissue cell while the other stays as a stem cell (in G0 arrest - at least it stays in G0 arrest until it receives signals to enter the cell cycle again). If one looks at the different events that control whether a cell enters the cell cycle from G0 arrest, it is clear that there are a lot of complicated steps involving a lot of different proteins to make it happen. Very often it takes a signal coming from outside the cell; for example: estradiol can do this for any cell with an estrogen receptor. Some growth factors and cytokines produced during inflammation also can do this. Once the cycle is started by these external signals, then internal regulatory signals take over and the cycle continues until the cell divides.
Events which alter the cell division cycle are very important in carcinogenesis. * * * While this is probably more information than humans should be allowed to understand note that growth factors, inflammatory cytokines & estrogen can start the cell division thing “rolling”.
While we can ignore most of the control mechanisms in the diagram you will notice that the growth factors as well as estrogen work by interacting with a regulatory protein called Rb. This protein is the key to understanding how HPV works. In order to force a cell to divide (and thereby replicate the virus) HPV genes code for a few proteins that interfere with the cell cycle control mechanisms. One of them is called E7 and this protein interferes with Rb function. Rb normally prevents entry into cell cycle and when E7 complexes with Rb the cell will enter the G1 phase of the cell cycle. (OK, so a lot more things happen and it is a lot more complicated than this but the point is clear, if you interfere with cell-cycle control mechanisms, you can get inappropriate cell division.) The end result is that the HPV infected epithelial cells which would not normally divide will start dividing and pile up to produce a benign tumor (a wart). Only after an adaptive immune response to these infected cells occurs will cytolytic T cells attack and get rid of the virus-infected cells (and the wart then disappears).
The series of reactions pictured here happen inside of those cells which have receptors for the signaling proteins. Note that these factors lead to increased protein synthesis and activation of Rb -
If one looks at the different events that control whether a cell enters the cell cycle from G0 arrest, it is clear that there are a lot of complicated steps involving a lot of different proteins to make it happen. Very often it takes a signal coming from outside the cell; for example: estradiol can do this for any cell with an estrogen receptor. Some growth factors and cytokines produced during inflammation also can do this. Once the cycle is started by these external signals, then internal regulatory signals take over and the cycle continues until the cell divides. If the HPV infection happens in cells with receptors for some of the external cell-cycle signals (lets say - cervical cells which respond to estrogen) then the cell division rate can be faster than in those cells that do not respond to external signals. This can have a detrimental effect, ie. decreased time for repair of DNA damage in these cells. If any DNA damage exists while the cell is duplicating all of the genes during the S phase, then there is a very high likelihood of producing a gene mutation in the resulting “daughter” cells. One form of DNA damage is the binding of any chemical to a DNA base. If there is an unwanted chemical bound to the DNA base, then that base will not be recognized as the correct base during DNA replication and a wrong DNA base will be put in its place instead. The end result is a change in DNA sequence. For example, if a hydroxyl radical (the most reactive and damaging of the oxygen radicals) binds to a Guanine then very often the G will be misread as an Adenine; resulting in a mutation (an A is there instead of the G). On the other hand, if a malondiadehyde (a larger carcinogen molecule produced during lipid peroxidation) binds to the G then a Thymine will be put there instead. The faster the rate of cell division, the less time for repair and the greater the likelihood of collecting mutations. If mutations occur in those proteins involved in controlling the cell cycle, then there is a high likelihood of the mutations leading to cancer (uncontrolled cell division). This progression towards cancer (collecting mutations with every cell division cycle until you mutate enough of the “right” genes to produce a cancer) occurs in cervical cells which are infected with HPV. One of the HPV genes also codes for another protein which is very much involved in this progression to cancer. One of the normal cell-cycle control proteins is called P53.
Also, please note that P53 is important for initiating transcription of proteins responsible for: • Shutting down the cell cycle when there is too much DNA damage • DNA repair • Initiating apoptosis when there is too much DNA damage to repair (next slide) *
P53 is an important transcription activator for many proteins; including the Fas & BAX proteins which are responsible for initiating apoptosis. P53 also inhibits synthesis of bcl2 which is a protein that blocks apoptosis. The end result is cells with elevated P53 will be killed.
P53 coordinates the stopping of the cell cycle and the repair of DNA damage when it exists. P53 does this by acting as an transcription activator to enhance synthesis of DNA repair enzymes (by inducing GADD45) and synthesis of P21, a protein which complexes with various proteins involved in the G1 phase to prevent further progression of the cell cycle until the DNA damage is repaired. Once the damage is repaired, then the cycle continues. Another important function of the P53 protein is to promote synthesis of proteins which activate the process of apoptosis when there is too much DNA damage to repair. Apoptosis is a process of cell death where the cell synthesizes an array of DNA’ase enzymes which then degrade genomic DNA to small fragments in order to destroy the cell. This is a form of cell suicide in order to protect the organism from developing cancer (or any other aberrant and possibly deadly form of mutation-induced cellular dysfunction).
So – what does this have to do with HPV? The HPV virus has genes which code for several proteins necessary for viral replication. Two of the proteins are important for the symptoms of HPV infection: E6 and E7. E6 binds to P53 and inactivates it E7 binds to Rb and inactivates it Because of these effects rates of cell division in infected cells will increase, rates of DNA repair will decrease, and rates of apoptosis will decrease.
Infection of epithelial cells with HPV will increase rates of cell division. Because skin epithelial cells do not respond to estrogen, there is a slight increase in cell division leading to the accumulation of cells; producing a benign tumor called a wart.
Infection of epithelial cells that are capable of responding to estrogen will greatly increase risk for cancer. There is a large increase in rates of cell division due to the combined effects of E6, E7, and Estrogen. Infected cells divide too fast for the DNA damage to be repaired and mutations accumulate, leading to increased risk for cancer P53 is partially inactivated which decreases rates of DNA repair, leading to the accumulation of more mutations and an increased risk for cancer Rates of apoptosis are reduced which increases the number of viable cells which contain DNA mutations, leading to an increased risk for cancer
Obviously, the E6 and E7 viral proteins are a major cause of increased risk for cancer as a result of HPV infection. Endogenous production of signals which can stimulate entry of stem cells or committed stem cells into the cell cycle (estrogen, inflammatory cytokines) are also intimately involved in the elevated risk for cancer resulting from HPV infection. Of the many different strains of HPV, several are resistant to immune recognition and infection with these strains results in long-term chronic infections while infection with those that are easily recognized results in relatively short-term infections. High risk for HPV-induced cervical cancer is only associated with those HPV strains which are resistant to immune recognition; indicating the possible importance of chronic inflammation in producing cancer risk. Thus, the end result of some HPV infections in cervical epithelia can be cervical cancer; although cervical cancer is relatively rare in HPV infected women, over 80% of cervical cancer cases are in HPV-infected women.
Infection by HPV can interfere with P53 function and the HPV protein called E6 is the problem here. This protein inhibits the function of P53, greatly increasing the likelihood that DNA damage will not be repaired, increasing the possibility that cells with too much DNA damage will NOT be killed, and therefore increasing the likelihood that mutations will occur. One reason why warts do not become cancerous is that skin epithelial cells do not normally divide and the rate of division in infected cells is quite slow. Cervical cells normally can divide so the rate of division in infected cells would be much faster, greatly increasing the risk for producing a cancer. Thus HPV infection = high risk for cervical cancer (and penile cancer too so don’t think the guys are “home free”) but not “wart” cancer. Now, just imagine if the P53 gene is one of the genes which acquire a mutation as a result of this process! Not only will its function be inhibited by the E6 protein but it will be destroyed by the mutation. Obviously this will not just increase risk for cancer but rather it will GUARANTEE cancer. One other source of risk from HPV infections which contributes greatly to the cancer process is that derived from the inflammatory process itself. Prolonged exposure of cells which are capable of dividing to the growth signals produced during an inflammatory response can ultimately lead to stimulation of cell division. A local infection which is resolved quickly is rarely relevant to the cancer process; however, a chronic infection which never resolves will lead to high risk for cancer. From a clinical standpoint, those strains of HPV which lead to warts or other skin/genital lesions which “go away” are rarely a risk for cancer. Those strains which are highly resistant to immune recognition and immune attack are the same strains which are associated with long-term chronic infection and high risk for cervical cancer. The introduction of the drug: Gardisil, is a major development for the prevention of HVP-caused cervical, penile, and anal cancer. HPV strains number 16, 18 (cervical cancer), 6, & 11 (genital warts) are the major causes of these two diseases while another 15 strains also are major risks for these same diseases. What the 19 HPV strains have in common is the inability of the human adaptive immune system to develop an effective response to their antigens, resulting in chronic infections. The vaccinations contain antigenic particles from the viruses that happen to be capable of being recognized by our adaptive immune cells and therefore will provoke an immune response against these viruses. The vaccination (prior to first infection) provides nearly 100% protection from both pre-cancer lesions and genital warts by insuring that on first infection the viruses will be killed.