Download
1 / 39
390 likes | 406 Views
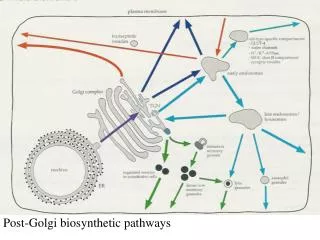
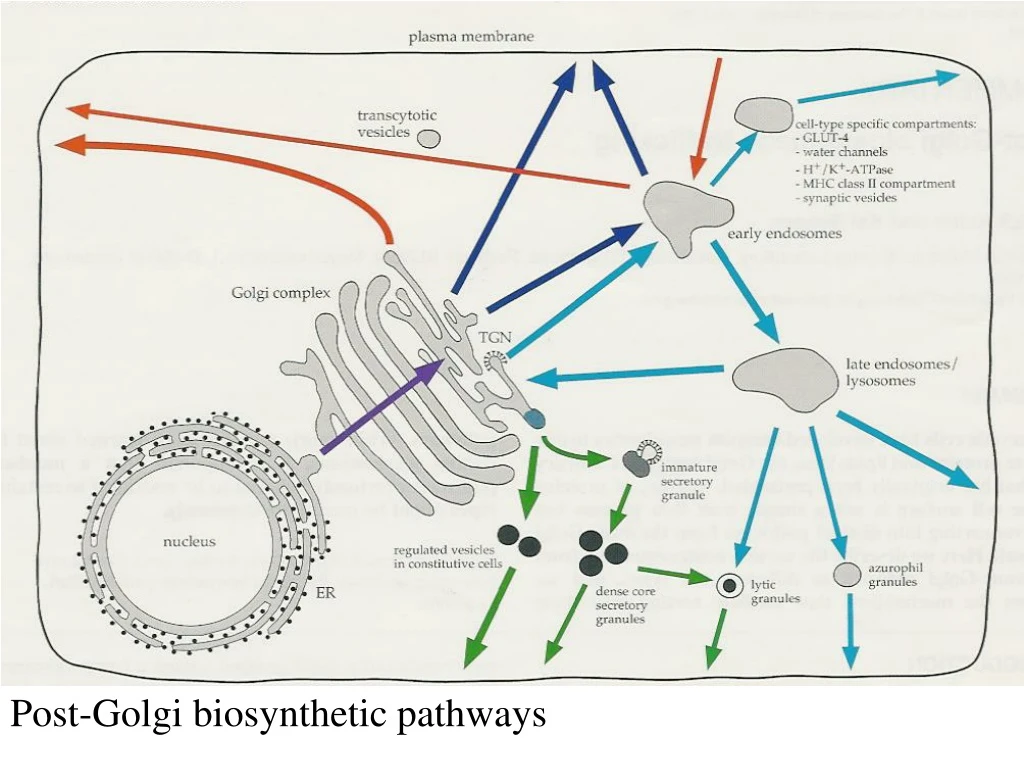
Post-Golgi biosynthetic pathways. MDCK-cell Resting fibroblast Migr ating fibrobl. The e pitel ial cell line MDCK is the most stud ied model system for polaris ed sort ing and transport. Hepatocyt e Retina l pigment Retina l rod cell

E N D
MDCK-cell Resting fibroblast Migrating fibrobl. The epitelial cell line MDCK is the most studied modelsystem for polarised sortingand transport.
Hepatocyte Retinal pigment Retinalrod cell epitelial cell
Hippocampus neuron Osteoclast Budding yeast cell
Sortingalong the biosynthetic pathway in epitelial cells. MDCK-cellsas modelsystem. Sorting in the trans-Golgi network. THE SORTING DEPENDSON SIGNALS IN THE MOLECULESTO BE SORTED
Classical signals: *Sortingof lysosomal enzymes to lysosomes *Basolateral transport in epithelial cells. *Retrograde transport from the Golgi to ER. *Endocytosisof receptorsandother molecules fromthe cell surface – a fraction is sorted to the trans-Golgi network.
Before any basolateral sortingsignalswere identified (1991 ->), it wassuggested that basolateral transport occured by “bulk flow” while transport tothe apicale side – which isthe specialiseddomain in epithelial cells – would require sorting. In 1991 it was published, however, that the transmembrane protein pIgA receptorwas transported basolaterallyin a signal dependent manner.
Growth of MDCK epitelial cellson filters. Transfer to glass-disheswith 90 ml of medium for establishmentof confluent cell layers.
Apikalt medium Basolateralt medium
PROTEIN SORTING STUDIES IN THE SECRETORY PATHWAY OF EPITHELIAL CELLS A protein transported equally well to the apical or basolateral surfaces is regarded as a protein without a sorting signal. Secretory proteins are the ones most easily studied, because these may be identified after harvest of the two opposite media of filter-grown cells. Apical or basolateral membrane proteins may be extracted after differetial biotinylation of the two opposite sides and/or recognition by specific antibodies. Sorting signals are autonomous – they work after ”transplantation” to normally non-sorted molecules.
Cytoplasmic domain Baso Api Baso Api Api Api Baso Api Lumenal domain tm
Lumen Lumenal domain Transmembrane domain Golgi membrane /endosomal membrane Cytoplasm Cytoplasmic domain
Truncated variant Lumenal domain TM domain Golgi membrane /endosomal membrane Apical Cytoplasmic domain Basolateral Example: Polymeric IgA receptor = pIgR
Truncated variant Lumenal domain TM domain Golgi membrane /endosomal membrane 3 aa Apical Cytoplasmic domain Basolateral Example: Polymeric IgA receptor = pIgR
Recessive apical sorting information Lumenal domain TM domain Golgi membrane /endosomal membrane Apical Cytoplasmic domain Basolateral Example: Polymeric IgA receptor = pIgR
Lumenal domain TM domain Golgi membrane /endosomal membrane 17 aa Basolateral Cytoplasmic domain Basolateral Apical Deletion of amino acids 655-668 = 14 amino acids
Lumenal domain TM domain Golgi membrane /endosomal membrane Baso Cytoplasmic domain Basolateral Apical 17 amino acids: RARHRRNVDRVSIGSYR, red identified by ala scan
Lumenal domain TM domain Golgi membrane /endosomal membrane Baso Apical Apical Reporter protein: PLAP (placental alkaline phosphatase) extracellular (lumenal) domain
WHAT MOTIFS ARE BASOLATERAL SORTING SIGNALS? YXX (YVEL/YTDI/YXRF) bulky/hydrophobic TGN38, M6PR LL / IL / LEL Fc Receptor, MHC II Invariant chain, EGF receptor NPXY Low density lipoprotein (LDL) receptor H/R-XXV Polymeric IgA receptor, Mannose-6-phosphate rec. (?) PXXP Epidermal growth factor (EGF) receptor Other Transferrin receptor (VDGDNSHVEMKLA) Some proteins have several sorting signals in their cytoplasmic tail. These are recognized at different intracellular sites, to provide recycling to the basolateral domain.
BASOLATERAL SORTING SIGNALS Some basolateral sorting signals overlap with endocytosis signals. *Fc receptor *Asialoglycoprotein receptor *Lysosomal acid phosphatase Other basolateral sorting signals are distinct from endocytosis signals. *Polymeric IgA receptor *LDL receptor *Transferrin recptor (?)
HOW AND WHERE IS BASOLATERAL SORTING INFORMATION RECOGNIZED? Adaptor complexes 1-3 Lysosomal route Basolateral route via endosomes
The first proteinsregarded as mediatorsof basolateral sorting were adaptins – already known to be involved in endocytosis from clathrin coated pits at the cell surface. The adaptins consist of 2 large, 1 medium and 2 small subunits. 4 different adaptin-complexes have been discovered. Baso A/B Baso AP-1A: TGN (+ endosomes) AP-1B: Epithelia specific AP-3A: Endosome/TGN AP-3B: Neuron-specific (endosome?)
AP-1, AP-2 and maybe AP-3 (inmammals) may bind clathrin. All 4 complexesare found inArabidopsis, but only AP 1-3 inDrosophila. Many of the subunits are found as closely related isoformes coded by separate genesmaking a large number of combinations possible. Endocytosis
AP-1B contains a specific1B subunitwhich only is expressed in certain polarised cells (not all polarised cell types, mainly epithelia). Recognizestyr-based signals. AP-4 has also been connected to basolateral sorting, but has equal or overlapping specificitywith AP-1B. There is still room formore adaptors for basolateral sorting. FAPP2 AP-4 Somewhere in the picture: FAPP1 and FAPP2, mediating TGN => PM transport.
GGA (1-3): Golgi-associated, -adaptin homologous, ARF-interacting proteins N-terminal hydrofobicsequence ARF-1 GDP GEF Rabaptin 5 binding Ear = GAE Ubq -synergin? What about tyr-signals? AP-1
Dynamin Cortactin Arp 2/3 complex Actin Membrane
At least 6 ARFs exist in mammals, 5 are localised to the Golgi-apparatusand 1 to the plasmamembrane. Aktiv membranbundet ARF1-GTP (myristoylated), a tyrosine based signal, andphosfatidylinositol 4,5 bisphosphatearenecessaryto recruit AP-1 clathrin adaptors to membranes. cytoplasmatisk 4 families GEFs with several members Phosphatidylinositidesof the 4-series hasbeen regarded as important for Golgi.
GGA dependent receptors CI-Mannose-6-phosphate receptor CD-Mannose-6-phosphate receptor Sortilin SorLA/LR11 LRP-3 -secretase
Some transmembrane proteins have cytoplasmic domains that interact directly with microtubule motors Non-clathrin dependent Clathrin-dependent
What about APICAL SORTING IN EPITHELIAL CELLS? Glycans: N-glycans, O-glycans, CS glycosaminoglycans Yes (maybe and no), yes (maybe), yes. GPI-anchors? NOT REALLY – OR? LIPID DOMAINS?? Protein motifs for apical sorting: Megalin NPXY. The second of three NPXY motifs is crucial (distance from membrane…).
MDCK cells transfected with the gene for the non-glycosylated protein rat growth hormone (rGH) secretes this protein randomly, which is slightly more basolaterally rGH with 2 N-linked glycosylation sites is secreted almost exclusively into the apical medium.
Erythropoietin – three N-glycans, one is critical. Endolyn – eight N-glycans, not all equally important. O-glycans of mucin type may also mediate apical sorting: Intestinal sucrase-isomaltase Gp-40 Several other examples But several examples of non-sorted glycoproteins also exist.
CS HS
Detergent insoluble proteins of apical transport vesicles were separated by 2D-gel analysis and sequenced. One putative lectin molecule was found: VIP 36. Hypothesis VIP 36? A raft Transmembrane lectin-molecule
WHATISA (GLYCOLIPID) RAFT? Glycolipid- andcholesterol rich domains ina lipidmembraneare associated in a more stable structure than lipids are according to “the fluid mosaic model.” On the cell surface of a “regular” cell, these domainswill have a diameter of 60 - 100 nm. In specialised membranesmay larger areas of the plasmamembrane have raft-characteristics. Example: The apical membraneof epithelial cells (MDCK). Do lipidsand lipid-binding proteinsplay a role in sortingof moleculesthat are transported from the TGN tothe apical membrane?
Caveolins: Proteinswith affinity for specialised lipid-domains. Palmitoylation. Might be necessary for cell surface transport of GPI-anchored proteins. Not apical transport. GPI-anker: Glycosyl-phosphatidyl-inositol-anchor that might bind proteins toa membrane. For some time regarded as sorting signals for apical transport, since these proteins usually are localised to rafts. The apical sorting is most likely dependent on N-glycans (via transcytosis?). Glycosphingolipids: are glycolipids that are mainly transported to the apical side in MDCK-cells (from the TGN). Present in rafts richin cholesterol. MAL (VIP 17):A protein that seems to mediate apical sortingofseveral cargo proteins in MDCK-cells.
There are probably several independent transport mechanisms operating in parallell, both to the apical and to the basolateral side of MDCK epitelceller. The apical onesmay be raft-basedor not raft-based.
Protein kinase D Annexin II, Annexin XIIIb, FAPP1/2, MAL (VIP17)
All mechanisms for sorting from the TGN are not known *We haveonly discussed proteinswith one transmembrane domain, while many proteins span the membrane several times.These may also be sorted. How? *Some apical proteins, like megalin, havebeen reportedto have signals in the cytoplasmictail (interaction with motor proteins?) *Ubiquitinylation may shift the sorting from TGN to the plasmamembrane towards TGN to lysosomes (the vacuole in yeast). *Lipids may play a role in sorting in many ways. *What factors are necessary for budding, transport and fusion? (v- and t- SNARES, GTP-binding proteins, etc.)