Download
1 / 33
441 likes | 857 Views
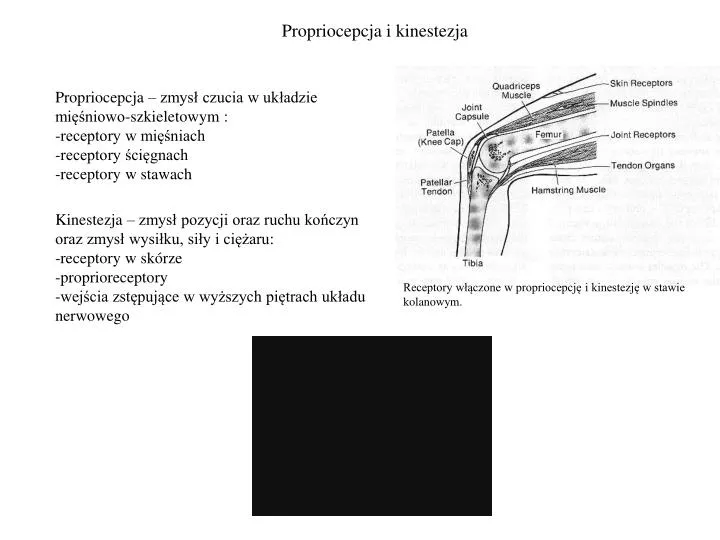
Propriocepcja i kinestezja. Propriocepcja – zmysł czucia w układzie mięśniowo-szkieletowym : receptory w mięśniach receptory ścięgnach receptory w stawach. Kinestezja – zmysł pozycji oraz ruchu kończyn oraz zmysł wysiłku, siły i ciężaru: -receptory w skórze -proprioreceptory

E N D
Propriocepcja i kinestezja • Propriocepcja – zmysł czucia w układzie mięśniowo-szkieletowym : • receptory w mięśniach • receptory ścięgnach • receptory w stawach Kinestezja – zmysł pozycji oraz ruchu kończyn oraz zmysł wysiłku, siły i ciężaru: -receptory w skórze -proprioreceptory -wejścia zstępujące w wyższych piętrach układu nerwowego Receptory włączone w propriocepcję i kinestezję w stawie kolanowym.
Ewolucja proprioreceptorów A. Brak zakończeń nerwowych w mięśniach. B. U płazów pojawiają się wrzecionka mięśniowe w mięśniach nóg. C. U ptaków i ssaków pojawiają się dwa rodzaje receptorów mięśniowych – z torebką jąder i z łańcuszkiem jąder. Dostają one osobne wejści motoryczne włóknami g.Pojawiają się unerwienia I-rzędowe (wychodzące ze środka wrzeciona) i II-rzędowe (wychodzące z peryferii wrzeciona). Pojawiają się narządy ścięgniste Golgiego w ścięgnach, służące do pomiaru napięcia mięśnia. Extrafusal – włókna ekstrafuzalne tworzące mięśnie, intrafusal – włókna intrafuzalne – są to zmodyfikowane włókna mięśniowe tworzące wrzecionka mięśniowe.
Wrzecionko mięśniowe A, B Układ eksperymentalny Ottoson, 1971: pojedyncze wrzecionko mięśniowe żaby wraz z pojedynczym wyjściem aferentnym, zamocowane pomiędzy dwoma nylonowymi nićmi. C. Zakończenia włókien motorycznych i sensorycznych w wrzecionku mięśniowym. Zakończenia włókien sensorycznych składają się z pęcherzyków, gdzie zachodzi transdukcja poprzez mechanicznie bramkowane kanały.
Odpowiedź wrzecionka mięśniowego Doświadczenia wykonane w układzie eksperymentalnym Ottosona. A. Odpowiedzi wrzecionka na różne prędkości rozciągania. B. Potencjał receptorowy (impulsy zablokowane farmakologicznie) dla różnych wartości rozciągnięcia. C.Częstość impulsów we włóknie aferentnym dla dynamicznego maksimum i statycznego ‘plateau’ odpowiedzi. D. Amplitudy potencjałów receptorowych dla fazy dynamicznej i statycznej rozciągania. Wniosek: wrzecionko mięśniowe precyzyjnie przekazuje informacje o szybkości zmian oraz stałym rozciąganiu mięśnia.
Odpowiedzi wrzecionka mięśniowego A. Rozciąganie mięśnia pasywnego (nie dostającego pobudzenia motorycznego). Szybko adaptacyjna odpowiedź z włókna I rzędowego i wolnoadaptacyjna z włókna II rzędowego. Włókna II rzędowe przenoszą informacje o pozycji. Narząd ścięgnisty wykazuje wysoki próg i małą czułość na rozciąganie mięśnia pasywnego. B. Rozciąganie mięśnia pasywnego z pobudzeniem motorycznym mięśni intrafuzalnych. Skurcz mięśni intrafuzalnych powoduje wzrost czułości na rozciąganie we włóknach I i II lecz nie wpływa na czułość narządu ścięgnistego. C. Krótkotrwały skurcz - rozkurcz mięśnia. Zwiotczenie mięśni intrafuzalnych powoduje brak impulsów we włóknach I i II. Narząd ścięgnisty wykazuje duża czułość podczas skurczu. D. Krótkotrwały skurcz - rozkurcz mięśnia z pobudzeniem motorycznym mięśni intrafuzalnych. W wyniku pobudzenia włóknami gamma, włókna intrafuzalnych nie wiotczeją w czasie skurczu i sygalizują stopień rozciągnięcia. Włókna gamma kontrolują niezależnie odpowiedzi dynamiczne (torebki) i statyczne (łańcuszki) zwiększając dokładność informacji sensorycznej.
Czucie w stawie A. Cztery rodzaje receptorów w stawie kolanowym. Typ I – analogiczne do zakończeń Ruffiniego w skórze (wolna adaptacja), Typ II - analogiczne do korpusku Paciniego w skórze (szybka adaptacja), Typ III - analogiczne do narządu ścięgnistego Golgiego. Typ IV – wolne zakończenia nerwowe analogiczne do zakończeń występujących w skórze. B. Odpowiedź receptora typu I u kota. Odpowiadają one tylko na pozycje w wąskich przedziałach kątowych na końcach skali. Sygnały z receptorów mięśniowych (linia przerywana) z mięśni przykolanowych współdziałąją w określaniu pozycji stawu.
Drogi wstępujące Droga rdzeniowo - móżdżkowa (lemniscal pathway) - dotyk i propriocepcja
Dwa rodzaje receptorów w układzie przedsionkowym II. Narządy otolitowe: łagiewka i woreczek – sygnalizują przyspieszenie liniowe I. Przewody półkoliste - sygnalizują przyśpieszenie kątowe
Rzęski i łączniki Rzęski komórek włoskowatych tworzą pęczek na szczycie komórki. Długość rzęsek nie jest jednakowa i zmienia się stopniowo. Na zdjęciu po prawej stronie widoczne są włókna (tip link) łączące końce sąsiednich rzęsek. Mają one średnice 3 nm i są pokryte metaliczną osłoną w celu wizualizacji. (ZAssad et al., 1991and Hudspeth and Gillespie 1944).
Model mechanizmu transdukcji w komórkach włoskowatych Kanały jonowe biorące udział w mechanoelektrycznej transdukcji w komórkach włoskowatych są bramkowane elastycznymi włóknami łączącymi.Otwieranie i zamykanie kanałów kontrolowane jest przez napięcie włókna, zależne od wychylenia pęczku rzęsek. Gdy pęczek rzęsek jest w spoczynku kanały przechodzą pomiędzy stanem otwartym i zamkniętym, spędzając większość czasu w stanie zamkniętym. Odchylenie pęczka w kierunku dodatnim zwiększa napięcie elastycznego włókna łączącego i prowadzi do otwarcia kanału i depolaryzcji komórki. Potencjał receptorowy zależy od kierunku wychylenia pęczka. Wychylenie w kierunku najdłuższej rzęski powoduje depolaryzacje i zwiększa częstość odpalania we włóknie wychodzącym. Wychylenie w kierunku przeciwnym powoduje hiperpolaryzacje i zmniejsza ilość wychodzących impulsów.
Mechanizm transdukcji Mechanotransdukcja w komórkach włoskowatych. A. Mechaniczna aktywacja kanału. Otwarcie następuje w wyniku naciągnięcia sprężyny. C. Łącznik odpowiada bramkującej sprężynie. Ugięcie w prawo powoduje napięcie łącznika, otwarcie mechanicznie czułych kanałów K+ i napływ jonów K+. Ugięcie w lewo hiperpolaryzuje komórkę włoskowatą.
Mechanizm transdukcji - adaptacja Motor – miozyna poruszająca się wzdłuż aktyny. A. Zanikająca odpowiedź dla stałego pobudzenia. B. Krzywa aktywacji przesuwa się podczas adaptacji, tak by akomodować przesunięcie włosków. C. Model ruchomego punktu zawieszenia. Motor zawsze wędruje do góry dopóki napięcie łącznika nie ściągnie go w dół.
Mechanizm transdukcji w komórkach włoskowatych cd • Ruch włosków • Naciągnięcie łącznika • Otwarcie kanału K+ • Napływ jonów K+ i depolaryzacja kmórki. Uwolnienie neurotransmitera z synapsy • Pobudzenie zakończenia nerwu przedsionkowego
Struktura czuciowa narządów otolitowych Ziarenka soli wapnia – otolity (kamyczki błędnikowe) Błona narządu otolitowego
Organizacja przestrzenna struktur czuciowych Woreczek -> płaszczyzna pionowa Łagiewka -> płaszczyzna pozioma Orientacja osi włosków w strukturze czuciowej narządów otolitowych (plamce). Wklęsła powierzchnia woreczka. Komórki włoskowate – czerwony, włoski – zielony, pozostałe komórki - niebieski
Dwa rodzaje komórek włosowatych Góra: dwa rodzaje komórek włoskowatych w grzebieniu bańkowym w kanale półkolistym u szynszyla. Dół: Nabłonek czuciowy w łagiewce. Dwa rodzaje komórek włoskowatych wyróżnione na podstawie unerwiających zakończeń synaptycznych. Włókna grube – duża czułość, szybka adaptacja. Włókna cienkie – niższa czułość, wolna adaptacja
Aktywność impulsowa we włóknach z kanałów półkolistych
Choroba Meniera • Choroba, której przyczyną jest zaburzenie wydzielania lub wchłaniania śródchłonki, co prowadzi do zwiększenia jej objętości. Choroba objawia się zawrotami głowy często związana z nudnościami, pogorszeniem słuchu, szumem i uczuciem pełności w uchu, zaburzeniami równowagi. • Zniszczenie błędnika może zapobiec zawrotom głowy ale nie ma wpływu na pozostałe dolegliwości. • Van Gogh mógł cierpieć na chorobę Meniera. Vincent van Gogh, Autoportret, 1889 I. K. Arenberg, L. F. Countryman, L. H. Bernstein and G. E. Shambaugh Jr,Van Gogh had Meniere's disease and not epilepsy, JAMA,Vol. 264 No. 4, July 25, 1990
Połączenia przedsionkowo - mózgowe Sygnały z komórek włoskowatych trafiają do jąder przedsionka leżących w pniu mózgu. W jądrach przedsionka zaczynają się trzy systemy projekcyjne: przedsionkowo – rdzeniowy (motoneurony kończyn, szyi i mięśni tułowia), przedsionkowo – oczny (motoneurony mięśni gałek ocznych), przedsionkowo – móżdżkowy. Drogi przedsionkowo – rdzeniowe dzielą się na drogi boczne i przyśrodkowe.
Boczne i przyśrodkowe obwody drogi przedsionkowo - rdzeniowej • Narządy otolitowe projektują głownie do dróg bocznych. Kanały półkoliste głownie do dróg przyśrodkowych. • Drogi boczne służą głownie, lecz nie wyłącznie, kontrolowaniu ruchu kończyn. Drogi przyśrodkowe kontrolują mięśnie szyi i tułowia. • Drogi boczne zawierają jedynie włókna pobudzające, drogi przyśrodkowe – zarówno pobudzające jak i hamujące. • W rdzeniu kręgowym drogi boczne tworzą połączenia poprzez interneurony (droga disynaptyczna), drogi przyśrodkowe tworzą bezpośrednie połączenia (droga monosynaptyczna). • Drogi przedsionkowe tworzą odruchowe przystosowania głowy i pozycji ciała w odpowiedzi na nachylenie i rotację.
Współpraca odruchów szyjnych i błędnikowych w odruchowym przystosowaniu kończyn w odpowiedzi na nachylenie ciała. Sygnały przedsionkowe (błędnikowe) określają położenie głowy, które zmienia się w wyniku nachylenia i obrotu głowy i ciała. Odruchy te działają na mięśnie szyi by przywrócić głowę do pozycji pozycji normalnej i na mięśnie kończyn by utrzymać równowagę Naciąg mięsni szyjnych wytwarza odruchowy skurcz mięśni szyjnych i kończyn. Odruchy przedsionkowe i szyjne mogą działać synergistycznie lub antagonistycznie
Odruch przedsionkowo – oczny: sygnały z kanałów półkolistych Obwody neuronalne odruchu przedsionkowo – ocznego. Informacja z kanałów półkolistych dociera do motoneuronów mięśni ocznych. Obrót głowy w lewo wywołuje ruch gałek ocznych w prawo, tak że obraz na siatkówce nie ulega przesunięciu Obrót głowy wywołuje ruch śródchłonki w przeciwnym kierunku w parze kanałów półkolistych. Wywołuje to zwiększenie aktywności jednego z kanałów i spadek aktywności drugiego.
Odruch przedsionkowo – oczny A. Ruch gałek ocznych jest kontrolowany przez sześć mięsni okołogałkowych. B. Zmiana spojrzenia bez udziału ruchu głową (np. czytanie). C. Zmiana spojrzenia oraz obrót głowy w kierunku patrzenia, tak by obiekt znajdował się w centrum pola widzenia. Fiksacja wzroku jest możliwa dzięki kompensacji ruchu oczu przez układ przedsionkowo - oczny
Odruch przedsionkowo – oczny cd Podczas utrzymującej się rotacji głowy gałki oczne ‘resetują’ spojrzenie tak, by obiekt znajdował się w centrum pola widzenia. Na zapisie widać pozycje oka podczas obrotu badanego ze stałą prędkością w ciemności. Ruch oczu składa się z fazy wolnej, o tej samej prędkości co obrót głowy, oraz ‘resetującej’ fazy szybkiej. Prędkość fazy wolnej stopniowo maleje aż do zaniku regularności. Konwergencja wzrokowych i przedsionkowych sygnałów na neuronach w neuronie przyśrodkowego jądra przedsionkowego. A. Obrót w ciemności – aktywność neuronu spada mimo utrzymywania się rotacji. B. Obrót w świetle - odpalanie neuronu jest stałe podczas rotacji. C. Zwierze jest unieruchomione, obraca się otoczenie. W stanie ustalonym neuron odpowiada tak jakby obracało się zwierzę lecz dłużej trwa dochodzenie do stanu ustalonego.
Stan nieważkości • Wrażenie raczej przyjemne • Szybka adaptacja(!) • Procesy adaptacyjne widoczne po powrocie na Ziemie, zanikające z czasem.