Download
1 / 56
560 likes | 672 Views
Remodeladores ATP dependientes. Remodeladores ATP dependientes.

E N D
Dynamic properties of nucleosomes. a | Remodelling complexes of the SWR1 family can remove the canonical H2A–H2B dimers and replace them with Htz1– H2B dimers (indicated in green), forming a variant nucleosome with unique tails that might bind unique regulatory proteins (Reg). b | Nucleosome modification (only acetylation (Ac) is depicted for simplicity) allows the binding of regulatory factors, which have specialized domains (bromodomains) that recognize acetylated histone tails. c | Nucleosome repositioning allows the binding of a regulatory factor to its site on nucleosomal DNA (light-blue segment). NATURE REVIEWS VOLUME 7 | JUNE 2006
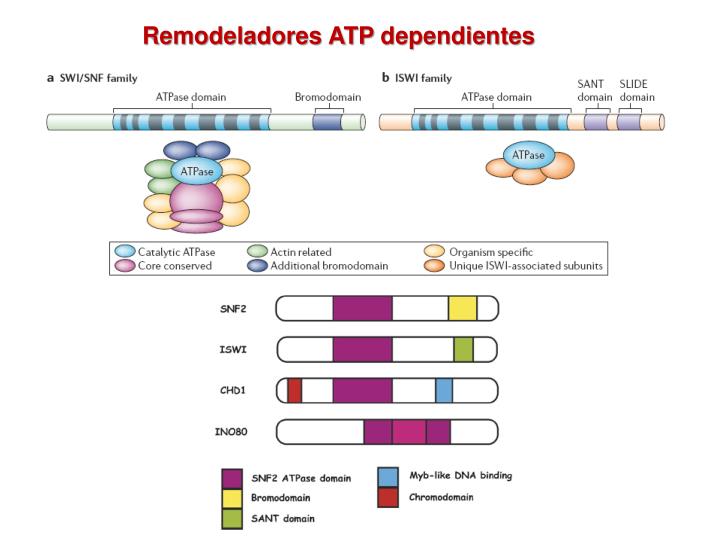
Properties de Sliding de los complejosremodeladores SWI/SNF y ISWI. NATURE REVIEWS VOLUME 7 | JUNE 2006
Figure 1. The Dynamic Nucleosome—A Substrate for Chromatin Remodeling ATPases Nucleosomes (shaded circles) are shown by looking down the axis of the 10 nm fiber. The 10 bp intervals at which DNA (blue line contacts the nucleosome are indicated by small red ovals. At the top is a schematic representing the propensity of the nucleosome to spontaneously unwrap and rewrap, starting at the entry and exit points (red arrow) for the nucleosomal DNA. The bottom portion is a schematic showing how capture of the intermediate unwrapped instate by an ATPase such as SWI/SNF may lead, by the loop-recapture model, to a 10 bp change in the translational rotation of thenucleonucleosome relative to the starting position on the DNA (yellow circles). For clarity, only one strand of DNA is shown. Molecular Cell, Vol. 19, 147–157, July 22, 2005
Figure 1. SWI/SNF-Dependent Stimulation of Transcription from Chromatin Templates
Figure 1 SWI/SNF displaces histones from DNA. a, Diagram of the substrate used in the sliding assay. b, The template does not enter the matrix of a polyacrylamide gel (lane 1) unless it has been digested with NheI, which releases the predominantly nucleosomal 39 end (lane 2). If the template is incubated with SWI/SNF before cutting, most of the 32P is released as freeDNA(lane 6) in an ATP dependent reaction (lane 5). In otherwise identical reactions, in which the substrate is cut before incubation with SWI/SNF, there is minimal histonedisplacement (lane 4). Nuc, NheI-digestednucleosomal DNA.
Figure 3 SWI/SNF-mediated transfer of octamer is blocked by a barrier in cis. a, Constructs containing four-way junction (4WJ) or control DNA inserts.
FIG. 1. AFM images of SWI/SNF and altered dimers. Samples were fixed, deposited, and imaged with a nanotube tip as described in Materials and Methods.
FIG. 3. 5S nucleosomal arrays are stably remodeled by hSWI/SNF. (A) Diagram of the 5S-G5E4 rDNA array used. White circles represent nucleosomes at preferred sites on 208-bp 5S rDNA sequences. Grey circles represent nucleosomes in the transcription template. Locations of SacI and XbaI sites relative to inferred unremodeled nucleosome positions are indicated. For the graphs, the array was treated with hSWI/SNF without ATP (triangles), with ATP for 30 min (squares), or with ATP for 30 min, followed by apyrase for 18 min (circles), and then cut with SacI (left) or XbaI (right) for the indicated times.The purified DNA was separated by agarose electrophoresis, and percent cutting was quantified
Modelo : traslocación del DNA , SWI/SNF NATURE REVIEWS. MOLECULAR CELL BIOLOGY (2006) 7 : 437-447
Figure 2. Dynamic Remodeling of a Nucleosome by an ATPase and a Transcription Factor (A) Periodicity and residence time of glucocorticoidreceptor (GR)-ligand (Dexamethasone[Dex]) interaction with promoter nucleosomes are determined by the amount of SWI/SNF (Brg) in thereaction. (B) Model summarizing the nucleosome dynamics during one cycle of GR-Dex/SWI/ SNF (Brg) interactions with promoter nucleosomes. Changes shown include changes to DNA-histone interactions (wavy lines) and the transient and sequential modifications to H2B (purple) and H2A (red) in the nucleosomes mobilized by SWI/SNF (Brg), followed by the cyclical collapse of the remodeled state into the ground state that correlates with displacement of GR/Dex and SWI/SNF. Taken from (Nagaich et al., 2004).
Mating-type switching FIGURE 1. Mating-type switching: switching from (a) MATa or (b) MATa. An HO endonuclease-induced double-strand break at MAT initiates gene conversion/replacement of the Ya region with Ya sequences copied from HMLa. MATa cells express Mata1p and Mata2p regulatory proteins, while MATa encodes Mata1p. HMLa and HMRa contain complete copies of mating-type genes but are not expressed because of the silencing imposed through the adjacent E and I silencer sequences that organize a repressed chromatin structure (indicated by hatched lines). HML shares more sequences (regions W, X, Z1 and Z2) with MAT than does HMR (regions X and Z1). MATa strains preferentially recombine with HMLa, even if HMR also contains Ya instead of Ya. Donor preference is dependent on a 244 bp cis-acting recombination enhancer (RE)
FT Swi/SNF SAGA
Cartoon of the assembled IFN-b enhanceosome. ATF-2/c-Jun binds to the PRDIV region. IRF-3 and IRF-7 bind to the PRDIII–I region and NFkB to the PRDII region. Atomic model of the INF-b enhanceosome (PDB file attached in the supplemental data section of the original article [6]). The p50 is in light blue and RelA in dark blue. IRF-7B and IRF-7D are in yellow and IRF-3A and IRF-3C are in green. ATF-2 is in red and c-Jun in blue. The DNA sequence is shown with the core-binding sites colored accordingly. Current Opinion in Structural Biology 2008, 18:236–242
PRDIV PRD III-I PRDII PRDIV PRDIII- I PRDII Figure 1. Comparison of the Specificity and Strengths of Synthetic and Natural Virus-Inducible Enhancers Comparison of the transcriptional activities of the intact lFN(3 promoter (bottom) with the individual homopolymeric PRDs after transient transfection into L929 cells. Each of the elements responds to virus infection as well as to other extracellular signals. By contrast, the lFN9 promoter is induced only by virus. Numbers indicate fold induction.
Figure 2. The Effects of Substitutions and Insertions on the Function of the IFNb Enhancer (A) Substitution of PRDIV by a second PRDII alters the specificity of induction. Human HeLa cells were transfected with the wild-type IFNb- CAT reporter construct or with the indicated PRDIV to PRDII substitution. The transfected cells were induced with either Sendai virus. The correct helical phasing of transcription factor-binding sites is required for virus mduction of the lFNf3 gene enhancer.
Figure 4. Premcubation of HMG l(Y) with the IFNB Enhancer Promotes the Cooperative Binding of IRF-I (A) Binding to the wild-type enhancer. Quantitative DNase I footprinting of the wild-type IFNp promoter is shown, using the same amounts of recombinant proteins detailed in Figure 3, except for IRF-1, which was 1, 2.5, 12, and 30 ng. In this experiment, HMG l(Y) (10 ng) was added to the DNA 10 min before the addition of the other proteins. (B) Binding to the enhancer containing a half-helical turn of DNA inserted between PRDI and PRDII. Quantitative DNase I footprinting
Modelo para la activación sinergística de la transcripción en el “enhanceosoma” del IFNb Figure 8. A Model for Synergistic Activation of Transcription by the IFNb Enhanceosome Virus infection leads to the cooperative assembly of the IFNb enhanceosome containing NF-kB (p50/p65 heterodimer), IRF1, andATF2/c-Jun. The high mobility group protein I(Y) that is required for enhanceosome assembly has been omitted from the picture for simplicity. The activation domains of each of the factors are indicated (AD) as distinct ovals. Due to the enhanceosome assembly, regions of the activation domains form a novel surface that constitutes a high-affinity binding site for the amino terminus of CBP as indicated. Recruitment of CBP by the enhanceosome tethers the polII holoenzyme via its interaction with the carboxyl terminus of CBP. Simultaneously, the activation domains of the activators contact the remaining components of the basal apparatus such as TFIID/A/B as indicated by the arrow. Shown is only a fraction of the components recruited and only some of the potential protein–protein interactions that lead to the assembly of the transcriptional machinery. Synergism in transcription results from the simultaneous recruitment of the two distinct subcomplexes, those of the polII holoenzyme and of the TFIID/A/B.
(D) Assembly of the IFN-b enhanceosome on nucleosomal IFN-b promoter fragments. An IFN-b promoter fragment (2143 to 1183) (lanes 1–7) or an identical-sized fragment bearing mutations in all HMG I(Y) binding sites (lanes 8–14) were reconstituted into a nucleosome, gel purified, and used in EMSA experiments along with recombinant IFN-b activators in the presence or in the absence of HMG
Figure 5. The IFN-b Enhanceosome Directs Ordered Recruitment of Histone acetyltransferases, SWI/SNF, and general transcription factors to the IFN-b promoter in vivo. The immunoprecipitations were repeated two times for p65, three times for hGCN5, three times for acetyl- Histone 4, three times for CBP, two times for PolII. two times for hBRG1, three times for TBP and two times for TAFII250. Shown are the results of one immunoprecipitation using all antibodies. The variability from experiment to experiment was small with respect to the kinetics of recruitment of individual factors.
Figure 6. Model Depicting the Ordered Recruitment of Chromatin-Modifying and Basal Factors to the IFN-b Promoter Shortly after virus infection, the enhanceosome assembles on the nucleosome-free IFN-b enhancer and recruits the GCN5 complex, which acetylates nucleosomes I and II (acetylated histone N-termini are shown as hooks). GCN5 departs from the promoter and the CBP-PolII holoenzyme complex is recruited by the enhanceosome. Next, SWI/SNF associates with the promoter via its interactions with CBP. This recruitment is stabilized by the acetylated histone N-termini, which presumably interact with the bromodomain of BRG1/BRM proteins present in the SWI/SNF complex. SWI/SNF remodels the nucleosomes (DNA shown as ruffled lines), thus allowing recruitment of TFIID, completion of preinitiation complex assembly at the core promoter and initiation of transcription (arrow).
Figure 1. Virus Infection Induces a Distinct Pattern of Histone Acetylation at the IFN- Promoter In Vivo (A) HeLa cells were either mock- or virus-infected with Sendai virus for the indicated amounts of time. Cross-linked chromatin was immunoprecipitated with the indicated antibodies and the IFN- promoter was detected by PCR in these samples using promoter-specific primers. The bottom part of the figure shows the abundance of the IFN- mRNA as detected by RT-PCR.
Schematic diagrams illustrating the modular structure of eukaryotic transcription activators.
HOMEOBOX Helix I Helix II Helix III
The leucine zipper, crystal GCN4 Dominiobásico y Leucine zipper
RECEPTORES NUCLEARES multi zinc fingers, dominio de union al DNA
Dominios de dimerización Leucine zipper Hélice: posiciónes g y e’ atracción electrostática Posiciones a-d y d-a’ interacciones hidrofóbicas