Download
1 / 54
550 likes | 761 Views
2 ATP + 4 ATP = + 2 ATP. So does this solve the direction problem? Only for a second … Where does this ATP come from, if we are E. coli growing in minimal medium… Glucose is the only carbon source. Need to make ATP from glucose, and this TAKES energy. Need only to regenerate ATP from ADP:.

E N D
2 ATP • + 4 ATP • = + 2 ATP
So does this solve the direction problem? Only for a second … • Where does this ATP come from, if we are E. coli growing in minimal medium… • Glucose is the only carbon source. • Need to make ATP from glucose, and this TAKES energy. • Need only to regenerate ATP from ADP: Via GLYCOLYSIS, e.g.
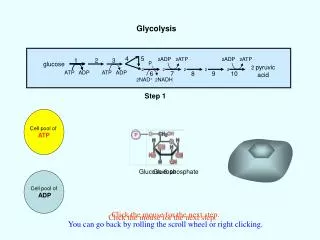
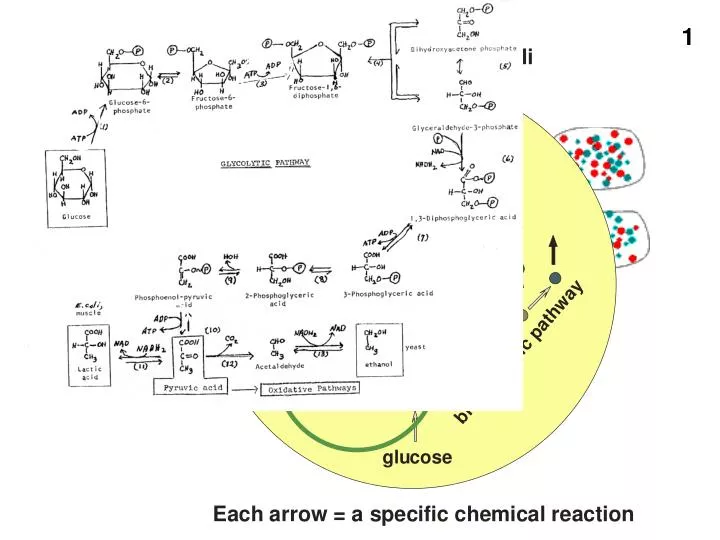
Overall reaction of glycolysis to pyruvate INCLUDING the generation of ATP 1 glucose + 2 ADP + 2 Pi + 2 NAD 2 pyruvate + 2 ATP + 2 NADH2 ΔGo = -18 kcal/mole So overall reaction goes essentially completely to the right.
The second way the cell gets a reaction to go in the desired direction: 1) A coupled reaction. One of two ways the cell solves the problem of getting a reaction to go in the desired directionGlucose + ATP glucose-6-P04 + ADP, ΔGo = -3.4 kcal/mole 2) The second way: • Removal of the product of an energetically unfavorable reaction • Uses a favorable downstream reaction • “Pulls” the unfavorable reaction • Operates on the second term of the ΔG equation. • ΔG = ΔGo + RTln([products]/[reactants])
So glucose pyruvic acid • ADP ATP, as long as we have plenty of glucose • Are we all set? • No…. What about the NAD.. We left it burdened with those electrons. • Soon all of the NAD will be in the form of NADH2 • Glycolysis will screech to a halt !! • Need an oxidizing agent in plentiful supply to keep taking those electron off the NADH2, to regenerate NAD so we can continue to run glucose through the glycolytic pathway.
Oxidizing agents around for NAD: 1) Oxygen Defer 2) Pyruvate In E. coli, humans: Pyruvate lactate, NADH2 NAD, coupled In Yeast: Pyruvate ethanol + CO2
Glucose excreted Handout 7-1b
Fermentation: anaerobiosis (no oxygen) Lactate fermentation Ethanolic fermentation Mutually exclusive, depends on organism Other types, less common fermentations, exist • (e.g., propionic acid fermentation, going on in Swiss cheese)
The efficiency of fermentation glucose--> 2 lactates, without considering the couplings for the formation of ATP's (no energy harnessing): ΔGo = -45 kcal/moleSo 45 kcal/mole to work with. Out of this comes 2 ATPs, worth 14 kcal/mol. So the efficiency is about 14/45 = ~30% Where did the other 31/45 kcal/mole go? Wasted as HEAT.
Fermentation goes all the way to the right glucose--> 2 lactates, without considering the couplings for the formation of ATP's (no energy harnessing): ΔGo = -45 kcal/mole kcal/moleOut of this comes 2 ATPs, worth 14 kcal/mol. So the efficiency is about 14/45 = ~30% Since 2 ATPs ARE produced, taking them into account, for the reaction: Glucose + 2 ADP + 2 Pi 2 lactate + 2 ATP ΔGo = -31 kcal/mole (45-14) Very favorable. All the way to the right. Keep bringing in glucose, keep spewing out lactate, Make all the ATP you want. That’s fermentation, for now.
Energy yield But all this spewing turns out to be wasteful. Glucose could be completely oxidized, to: … CO2 That is, burned. Complete oxidation of glucose, Much more ATP But nature’s solution is a bit complicated. The fate of pyruvate is now different How much energy released then? Glucose + 6 O2 6 CO2 + 6 H2O ΔGo = -686kcal/mole ! Compared to -45 to lactate (both w/o/ ATP considered)
Acetyl-CoA Score: Per glucose 2 NADH 2 NADH 2 ATP 2 CO2
Acetyl-CoA O || CH3 - C –OH + Co-enzyme A Acetyl ~CoA Acetic acid, acetate Acetate group
Acetyl-CoA Per glucose 2 oxaloacetate 2 NADH2 NADH 2 NADH 2 NADH 2 ATP 2 CO2 2 CO2 2 CO2 6 CO2
GTP is energetically equivalent to ATP GTP + ADP GDP + ATP ΔGo = ~0 G= guanine (instead of adenine in ATP)
Acetyl-CoA Per glucose 2 oxaloacetate 2 NADH 2 NADH 2 NADH 2 NADH 2 ATP 2 ATP 2 CO2 2 CO2 2 CO2 2 Succinic dehydrogenase
FAD = flavin adenine dinucleotide FAD + 2H. FADH2
Acetyl-CoA Per glucose oxaloacetate 2 NADH 2 NADH 2 NADH 2 NADH 2 FADH2 2 NADH 2 ATP 2 ATP 2 CO2 2 CO2 2 CO2 Succinic dehydrogenase
Per glucose 2 NADH 2 NADH 2 NADH 2 NADH 2 FADH2 2 NADH 2 ATP 2 ATP Glucose + 6 O2 6 CO2 + 6 H2O : By glycolysis plus one turn of the Krebs Cycle: 1 glucose (6C) 2 pyruvate (3C) 6 CO2 2 X 5 NADH2 and 2 X 1 FADH2 produced per glucose 4 ATPs per glucose NADH2 and FADH2 still must be reoxidized …. No oxygen yet to be consumed No water produced yet 2 CO2 2 CO2 2 CO2
Oxidation of NAD by O2 NADH2 + 1/2 O2 --> NAD + H2O ΔGo= -53 kcal/mole If coupled directly to ADP ATP (7 kcal cost),46 kcal/mole waste, and heat So the electrons on NADH (and FADH2) are not passed directly to oxygen, but to intermediate carriers, Each transfer step involves a smaller packet of free negative energy change (release)
NADH2 H Ubiquinone; Coenzyme Q H Handout 8-3
Schematic idea of H+ being pumped out nal Handout 8-4
FoF1 complex Handout 8-4
Chemiosmotic theory Proton motive force (pmf) Chemical gradient Electrical gradient Electrochemical gradient Peter Mitchell 1961 Water-pump-dam analogy Some evidence:
Artificial phospholipid membrane H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ ETC Complex I’s pH drops pH rises NADH NADH H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+
Artificially produced mitochondrial membrane vesicle ATP is formed from ADP + Pi
Dinitrophenol (DNP): an uncoupler of oxidative phosphorylation - + H+ DNP’s -OH is weakly acidic in this environment DNP can easily permeate the mitochondrial inner membrane Outside the mitochondrion, where the H+ concentration is high, DNP picks up a proton After diffusing inside, where the H+ concentration low, it gives up the proton. So it ferries protons from regions of high concentration to regions of low concentration, thus destroying the proton gradient. Electron transport chain goes merrily on and on, but no gradient is formed and no ATP is produced.
The mechanism of ATP formation: The ATP synthetase (or ATP synthase) The F0F1 complex
outside inside Gamma subunit: cam ATP synthetase
inside outside
Alpha+beta Gamma
Actin labeled By tagging it with fluorescent molecules Actin is a muscle protein polymer Testing the ATP synthetase motor model by running it in reverse (no H+ gradient, add ATP)
Run reaction in reverse, add ATP, drive counter-clockwise rotation of cam 4 3 2 1 5 ATP hydrolysis This is oxidative phosphorylation of ADP
Actin labeled By tagging it with fluorescent molecules Actin is a muscle protein polymer Testing the ATP synthetase motor model by running it in reverse
ATP accounting • Each of the 3 ETC complex (I, III, IV) pumps enough H+ ions to allow the formation of 1 ATP. • So 3 ATPs per pair of electrons passing through the full ETC. • So 3 ATPs per 1/2 O2 • So 3 ATPs per NADH2 • But only 2 ATPs per FADH2 (skips complex 1)
ATP ATP ATP Similar to handout 8-2
OXPHOS: 1 NADH from glycolysis 1 NADH from Krebs entry 3NADH from Krebs 1 FADH2 from Krebs Total: 17 ATP 5 NADH = 15 ATP 1 FADH2 = 2 ATP Substrate level phosphorylation (SLP): 2 ATP 1ATP from Glycolysis 1 ATP (GTP) from Krebs Handout 8-6 Grand total (E. coli): 17 + 2 = 19 per ½ glucose or 38 per 1 glucose