Download
1 / 116
1.16k likes | 1.27k Views
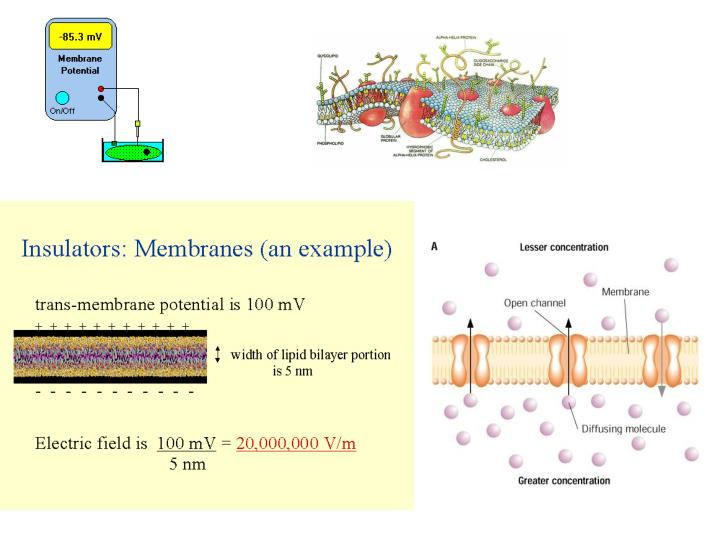
Little change in concentration. 외부의 자극에 의해 흥분되지 않은 resting 상태에서 그대로 일정한 membrane potential(MP) 을 유지 ---> resting potential(RP) nerve, muscle 에서 항상 negative, constant, cell type 에 특징적 e.g.: warm-blooded animal: -55 to -100 mV; smooth muscle: -30 mV

E N D
외부의 자극에 의해 흥분되지 않은 resting 상태에서 그대로 일정한membrane potential(MP)을 유지---> resting potential(RP) • nerve, muscle에서 항상 negative, constant, cell type에 특징적e.g.: warm-blooded animal: -55 to -100 mV; smooth muscle: -30 mV • 이러한 RP은 막의 채널을 통해 ion들이 passive하게 이동함으로서 발생한다. • Resting시 대부분의 열린 채널은 K+ ion, 따라서 RP은 주로 K+ ion에 대한 transmembrane concentration gradient에 의해 주로 결정된다.
A-: proteins, polypeptides, organophosphates (such as DNA, RNA, nucleotides such as ATP, and an array of nutrients. They all behave as organic acids, giving off a hydrogen ion (that is incorporated with oxygen to form metabolic water) and leaving the negative ion inside the cell. This establishes a negative charge dispersed throughout the cytoplasm, and positive ions are attracted.
Potassium equilibrium -90 mV
Nernst Equation By the end of the 19th century, it was known that the cytoplasm was high in K+ and that [Na+] was very low--and that this relationship was reversed outside the cell. The assumption was made that the cell membrane was permeable to K+ but not to Na+. Direct measurement of the transmembrane potential was not yet possible, but an effort was made to calculate this voltage using the Nernst equation, shown at left.
Nernst equation Eion is the “equilibrium potential” for a single permeant ion Eion(mV) = 58 / charge • log ([ion]out / [ion]in) E?+(mV) = –58 mV = 58 / +1 • log (10 mM/ 100 mM) EK+(mV) = –81 mV = 58 / +1 • log (5 mM/ 125 mM) ENa+(mV) = +58 mV = 58 / +1 • log (150 mM/ 15 mM) ECa+(mV) = +145 mV = 58/ +2 • log(100mM/ 0.001mM) ECl–(mV) = –58 mV = 58 / –1 • log ( 100 mM/ 10 mM)
Goldman equation was derived to solve for transmembrane potential using all ions involved simultaneously. For most animal cells, the only important ions are K+, Na+, and Cl-. Goldman includes the permiability factor, p, of each ion. Solution of the Goldman equation for a cell membrane yields an accurate model of the transmembrane voltage at any particular set of concentrations and temperatures. the inclusion of chloride ion concentrations in Goldman equations is redundant. Omit the choride ion, and you will get the same answer. Later, when you think about action potentials, you will realize that only the sodium and potassium ions migrate across the membrane in significant numbers, so only they are practically important in understanding membrane--ion interrelationships.
lessons: • • The resting potential is based on the fact that K+ is actively pumped into all cells. • Given the concentration gradients of K+, & the selective permeability of the resting to K+, the resting potential results by diffusion with no energy required. • Ions pass membrane through protein pores called channels. • Equilibrium is reached when there is a balance between tendency to diffuse & electrical attraction. • The membrane of cells is permeable to both K+ and Na+, so both ions contribute to the resting potential. • Na+ “leakage” into cell must be counter-acted with a Na/K exchange pump. • Electrical potential changes as current spreads along membranes. • Electrical properties of membrane determine how the potential changes over distance and over time.
Electroneutrality Channels Membrane potential Generation of a Membrane Potential ORIGIN OF THE NERNST POTENTIAL Membrane Potential Affects Ion Movement K+ channel subunit structureK+ channel, extracellular view, S5 and S6 segmentsVoltage Sensor: rest vs. activeK+ channel structure & K+ current measurementNa+ channel alpha subunit
Depolarization The animation below illustrates how the flow of positively charged ions into the axon leads the axon to become positively charged relative to the outside. This initial phase of the action potential is called the depolarization phase. At the end of the depolarization phase, the voltage of the inside of the axon relative to the outside is positive and the relative concentration of sodium ions inside the axon is greater than at the beginning of the action potential.
Trigeminal Ganglion Cell: this is about 2 seconds of activity that was recorded from a rat ganglion cell after a single whisker (vibrissa) was moved and held in position. This neuron was firing about 100 action potentials every second. Each action potential in this record is separated by about 10 milliseconds. There are 21 action potentials displayed in this picture of the recording - count them! Action potential recording
The action potential indicates what happens when the neuron transmits information from one cell to another. Neuroscientists use other words, such as a "spike" or an "impulse" to describe the action potential. The action potential is an explosion of electrical activity that is created by a depolarizing current. This means that some event (a stimulus) causes the resting potential to move toward 0 mV. When the depolarization reaches about -55 mV a neuron will fire an action potential. This is the threshold. If the neuron does not reach this critical threshold level, then no action potential will fire. Also, when the threshold level is reached, an action potential of a fixed sized will always fire...for any given neuron, the size of the action potential is always the same. There are no big or small action potentials in one nerve cell - all action potentials are the same size. Therefore, the neuron either does not reach the threshold or a complete action potential is fired - this is the "ALL OR NONE" principle.
The "cause" of the action potential is an exchange of ions across the neuron membrane. A stimulus first results in the opening of sodium channels. Since there are a lot more sodium ions on the outside, and the inside of the neuron is negative relative to the outside, sodium ions rush into the neuron. Remember, sodium has a positive charge, so the neuron becomes more positive and becomes depolarized. It takes longer for potassium channels to open. When they do open, potassium rushes out of the cell, reversing the depolarization. Also at about this time, sodium channels start to close. This causes the action potential to go back toward -70 mV (a repolarization). The action potential actually goes past -70 mV (a hyperpolarization) since the potassium channels stay open a bit too long. Gradually, the ion concentrations go back to resting levels and the cell returns to -70 mV.
Distribution of channels Leak channels everywhere
Voltage-gated Na+ channel Not “ready” But “ready” Pass current @ “threshold voltage”
Positive feedback loop graded Na+ potential If YES, then... Reach “threshold”? V-gate Na+ channels open Na+ enters (depolarization)
Action potential initiation S.I.Z.
Action potential termination Think “votes”
Voltage Clamp If resistance decreases and current stays the same, then voltage also decreases so that the result of V/R stays the same - I. So if R decreases and I is constant, V decreases. V is held constant while R is decreasing. If R decreases, then I must increase to keep V constant. So if V is constant while R is decreasing, then I must increase. If you can measure I and V, then R can be easily calculated by re-arranging the equation to: This is the same as question 2. If voltage is constant and resistance decreases, what happens to current? Current must increase if voltage remains constant when resistance deceases. You can see this in the figure on the right - ion channels open and positively charged ions can flow into the cell.
Sodium Conductance Activation and Inactivation Changes in sodium and potassium conductance with different depolarizations. activation & inactivation. When inactivation is present, sodium channels cannot open again. The peak conductance increases with greater depolarization. inactivation slowly goes away, eventually allowing sodium channels to open again if depolarization occurs. During the period of inactivation, it is more difficult to increase gNa, and therefore the presence of inactivation underlies refractoriness, i.e. the inability to trigger an action potential for some time after another action potential has occurred.
Voltage clamp analysis At membrane potentials above +50 mV sodium actually exits the nerve cell. At +50 mV there is no sodium flow timing of important events in the cell sodium influx is at a maximum at about 1 millisecond. Potassium efflux on the other hand is at a maximum at about 3 ms. TTX (tetrodotoxin) effectively blocks the sodium channels in nerve cells. A drug called TEA (tetraethylammonium) will effectively block potassium flow and allow for the testing of membrane conductance and timing for sodium influx.
Hodgkin-Huxley equations Hodgkin-Huxley equations describe the generation and propagation of the action potentials in neurons in terms of the dynamics of the potassium and sodium channels in the axon membrane. The conductance of the channels is described in terms of internal state variables of the channel (h, m and n below). The probability that a channel will open is a function of the state variables, the total conductance of a population channels is the product of the channel opening probability, the single channel conductance and the total number of channels. The Hodgkin-Huxley equations are: Where each of n, m and h satisfy equations of the type: Here w, z, , , Ileak, VNa and VK are experimentally determined constants, e and k are constants of nature, T is the temperature and and experimentally determined, voltage dependent, rate constants. An input current injection, Iinj may cause the membrane potential to exceed a threshold so that it starts firing action potentials. Action potentials last around 10ms, and the important time scale is about 10-100 s.
The animation to the left shows an idealized neuron. The red ovals represent Na+ ion channels and the green ovals represent K+ ion channels. You can start the movie and it will show an action potential (shown as a colored curve) move down the axon. When the sodium channels open during the depolarization (the red section of the action potential curve), the Na+ rushes in because both of the greater concentration of Na+ on the outside and the more positive voltage on the outside of the axon. When the Na+ channels close and the K+ channels open (the green section of the action potential curve), the K+ now leaves the axon due both to the greater concentration of K+ on the inside and the reversed voltage levels. Thus, in many ways the action potential is not the movement of voltage or ions but the flow of these ion channels opening and closing moving down the axon. This movement of the ion channels explains why the action potential is slow relative to the normal flow of electricity. The normal flow electricity is the flow of electrons and travels at the speed of light while these ion channels movement is considerably more slowly. These are mechanical movements and cannot move nearly at the speed of light.
Myelin wraps around a nerve cell axon in the spinal cord. In the CNS, glial cells called oligodendrocytes produce myelin, which is composed of multiple layers of oligodendrocyte membranes that wrap concentrically around one or more axons. (In the PNS, Schwann cells form myelin.) The myelin sheath around an axon acts like an electrical insulator, allowing nerve impulses to be conducted very quickly. Myelin appears white and shiny because the layers of membrane that form it contain large amounts of fatty substances called lipids.
The central nervous system consists of neurons and glial cells. Neurons constitue about half the volume of the CNS and glial cells make up the rest. Glial cells provide support and protection for neurons. They are thus known as the "supporting cells" of the nervous system. The four main functions of glial cells are: 1. to surround neurons and hold them in place, 2. to supply nutrients and oxygen to neurons, 3. to insulate one neuron from another, and 4. to destroy and remove the carcasses of dead neurons (clean up). The three types of CNS supporting cells are Astrocytes, Oligodendrocytes, and Microglia. The supporting cells of the PNS are known as Schwann Cells.