Download
1 / 121
1.22k likes | 1.4k Views
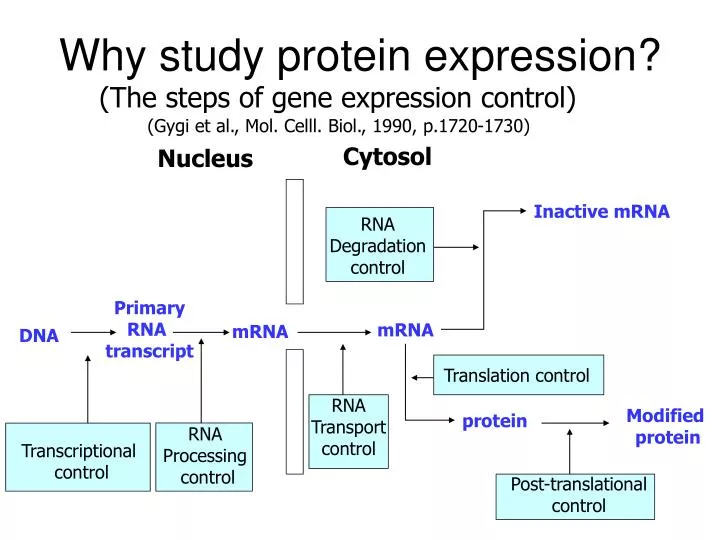
Inactive mRNA. RNA Degradation control. Primary RNA transcript. mRNA. mRNA. DNA. Translation control. RNA Transport control. Modified protein. protein. RNA Processing control. Transcriptional control. Post-translational control. Why study protein expression?.

E N D
Inactive mRNA RNA Degradation control Primary RNA transcript mRNA mRNA DNA Translation control RNA Transport control Modified protein protein RNA Processing control Transcriptional control Post-translational control Why study protein expression? (The steps of gene expression control) (Gygi et al., Mol. Celll. Biol., 1990, p.1720-1730) Cytosol Nucleus
真核生物基因表达的调控 10.1 染色质结构与基因表达 染色质是细胞核中基因组DNA与蛋白质构成的复合体。染色质的基本结构单位是核小体。10 nm粗的纤维可以进一步盘绕成30 nm粗的纤维。在分裂期,30 nm粗纤维再折叠成具有一定形态结构的染色体。分裂期结束后,染色体又转化为染色质。按照功能不同,可将染色质划分为活性染色质和非活性染色质。前者是指那些具有转录活性的染色质,而后者则用于表示缺乏转录活性的染色质。
在结构上,活性染色质和非活性染色质也有很大的差异。具有转录活性的染色质区域为一种开放、松散的结构。而非活性染色质呈现一种高度浓缩的形态,转录机器不能与其中的启动子结合,因而没有转录活性。异染色质就是一种典型的非活性染色质。在结构上,活性染色质和非活性染色质也有很大的差异。具有转录活性的染色质区域为一种开放、松散的结构。而非活性染色质呈现一种高度浓缩的形态,转录机器不能与其中的启动子结合,因而没有转录活性。异染色质就是一种典型的非活性染色质。
10.1.1 位置效应 位置效应(position effect)是指一个基因由于在基因组的位置发生改变,而发生的表达上的变化。位置效应在果蝇中得到了详尽的研究。野生型w+基因使果蝇的复眼呈红色。有一个果蝇品系,由于染色体倒位使w+基因座靠近着丝粒处的异染色质。
10.1.2 活性染色质的特征 与非表达区域中核小体结构紧密、间隔规则相比,活性染色质区域的核小体组装较为伸展或不规则。这样的一种结构有利于转录因子的结合,以及RNA聚合酶沿模板的滑动。在转录起始区以及某些特殊的区域,核小体的构象变化更为明显,DNase I和微球菌核酸酶等非特异性内切酶可用于检测这种变化。
在染色质中还存在一些短的对DNase I的消化十分敏感的区段,长度一般介于50~200 bp之间,可被极微量的DNase I降解,被称为DNase I超敏感位点(DNase I hypersensitivity site)。通过分析很多基因的染色质,发现DNase I超敏感位点广泛存在。每个活跃表达的基因都有一个或几个超敏感位点,大部分位于基因的5’调控区,少数位于其他部位,如转录单位的下游。非活性基因的5’侧翼区的对应位点不会表现出对DNase I的敏感性。
超敏感位点代表着开放的染色质区域,由于组蛋白八聚体的解离或缠绕方式的改变,这段区域中的DNA序列暴露,易受到核酸酶的攻击。对超敏感位点的最初认识来自SV40微小染色体的研究。SV40的DNA是5200 bp的环状分子,周长1500 nm。在宿主细胞的细胞核中,SV40形成串珠状核小体。在它的复制起点附近,同时也是晚期转录启动子的上游,存在一段DNase I的超敏感区域,在电镜下可直接观察到该区域在核小体组装上出现空缺。这段空缺长约120 nm(约350 bp),无核小体结构。
10.1.3 染色质结构的调节 在原核细胞中,RNA聚合酶和调节蛋白可以自由地接近DNA。在真核细胞中,由组蛋白和基因组DNA两部分组成的染色质结构限制了转录因子对DNA的接近与结合,实际上起着阻遏转录的作用。基因转录需要染色质发生一系列重要的变化,如染色质去凝集,核小体变成开放式的疏松结构,使转录因子等更容易接近并结合核小体DNA。有两种方式可以显著改变DNA的易接近性:组蛋白的乙酰化和核小体重塑。组蛋白的去乙酰化,则可以使染色质凝集,引起基因沉默。
10.1.3.1 组蛋白N端尾的修饰对染色质结构及基因转录的影响 核心组蛋白保守的折叠结构域和N端尾巴 Histone fold:~80个氨基酸残基构成的保守的区域由3个α螺旋组成,螺旋间由短的无规则的环隔开。
组蛋白尾含有的几个赖氨酸残基是乙酰化的位点。核心组蛋白赖氨酸的乙酰化中和了它的正电荷,降低了组蛋白尾对带负电荷的DNA骨架的亲和性,导致局部DNA与组蛋白八聚体解开缠绕,从而为参与转录调控的各种蛋白质因子与DNA的结合创造了条件。组蛋白N端尾对30 nm纤维的形成是必需的,当N端尾被乙酰化后,从核小体纤维盘绕成30 nm粗纤维的过程将被阻止。异染色质中的组蛋白一般不被乙酰化,而具有转录活性的染色质常常是高度乙酰化的,这些清楚地表明这种类型的修饰与DNA的包装相关。
经过多年的努力,催化向组蛋白添加乙酰基的组蛋白乙酰转移酶(histone acetyl transferase, HAT)终于在1996年被分离出来。许多组蛋白乙酰转移酶是以往鉴定过的激活蛋白或辅激活蛋白(coactivator)。
激活蛋白Gcn4募集辅激活蛋白Gcn5复合体指导启动子处组蛋白N端尾的乙酰化激活蛋白Gcn4募集辅激活蛋白Gcn5复合体指导启动子处组蛋白N端尾的乙酰化 四膜虫的P55蛋白质是最先发现的一种乙酰转移酶。
组蛋白乙酰化为一可逆过程,乙酰化和去乙酰化的动态平衡控制着染色质的结构和基因表达。组蛋白脱乙酰酶(histone deacetylase, HDAC)可去除组蛋白上的乙酰基,抑制基因表达。当第一个组蛋白去乙酰化酶从人类细胞中被分离出来后,组蛋白的去乙酰化与基因转录抑制之间的关系就建立起来了。
抑制子Ume6指导Sin3复合体的去磷酸化作用 Ume6:抑制子,阻遏蛋白
10.1.3.2 核小体重塑 核小体重塑涉及在基因组一个较短的区域中核小体位置的改变或者结构的改变,是一个能量依赖的过程,由核小体重塑复合体(chromatin remodeling complex)催化完成。重塑复合体利用ATP水解释放的能量,介导二种主要的反应:(1)组蛋白八聚体沿DNA滑动,从而改变核小体的位置,使转录因子能够与原来无法靠近的启动子序列结合;(2)介导组蛋白重排,在不改变位置的情况下使核小体变成一种较为松散的结构。
10.1.4 染色质结构的区间性 10.1.4.1 绝缘子 绝缘子(insulator)序列首先在果蝇中被发现,以后又在多种真核生物的基因组中被鉴定出来。1985年,Udvadry等在果蝇的基因组中检测出了两个绝缘子序列,scs和scs’(specialized chromatin structure)。scs和scs’分别位于果蝇多线染色体S7A7条带热激蛋白基因座的两侧,长度分别是350 bp和200 bp。当受到热激时,hsp70基因高水平转录,在多线染色体上形成一个膨泡。
scs和scs’就位于膨泡的两端,是膨泡的边界元件。在scs和scs’的外侧是异染色质区域,所以绝缘子能够抵挡异染色质对热激蛋白基因座位的影响,使该座位在结构和功能上都是一个独立的区域。scs和scs’就位于膨泡的两端,是膨泡的边界元件。在scs和scs’的外侧是异染色质区域,所以绝缘子能够抵挡异染色质对热激蛋白基因座位的影响,使该座位在结构和功能上都是一个独立的区域。 350 bp S7A7 200 bp
绝缘子是一组在真核生物基因组中建立独立的转录活性区的调控元件,它具有两种性质。第一,当绝缘子位于增强子或者沉默子与启动子之间时可以阻断它们对启动子的作用;第二,绝缘子可以使其界定的基因的表达不受位置效应的影响。在转基因时,目的基因整合到染色体上的不同位置,因位置效应作用基因的表达水平差异很大。但是,如果在目的基因的两侧连接上绝缘子,目的基因往往不受染色体位置效应的影响。绝缘子是一组在真核生物基因组中建立独立的转录活性区的调控元件,它具有两种性质。第一,当绝缘子位于增强子或者沉默子与启动子之间时可以阻断它们对启动子的作用;第二,绝缘子可以使其界定的基因的表达不受位置效应的影响。在转基因时,目的基因整合到染色体上的不同位置,因位置效应作用基因的表达水平差异很大。但是,如果在目的基因的两侧连接上绝缘子,目的基因往往不受染色体位置效应的影响。
10.1.4.2 基质附着区(matrix attachment region, MAR) 在细胞间期,由丝状蛋白构成的网络状的核基质附着于核膜的内表面。DNA借着于基质附着区(matrix attachment region,MAR)与基质蛋白结合。因为MAR也被用于附着染色体支架,因此也称为支架附着区(scaffold attachment region, SAR)。这些MAR/SAR位点长度为200-1000 bp,富含AT(占70%),但是没有明显的一致序列。具有几个连续腺嘌呤的DNA序列有发生弯曲的趋势。与MAR位点结合的核蛋白识别弯曲的DNA,而不是特定的序列。
10.1.4.3 基因座控制位点 人类的β-珠蛋白基因簇长约60 Kb,含有5个功能基因,排列顺序是5’- ε-γG-γA-δ-β-3’。其中ε在胚胎早期表达,γG和γA在胎儿时期表达,δ和β在成体中表达。每个β-珠蛋白基因都有自己的一套调控系统,但它们的表达还要受到基因簇上游12 Kb的基因座控制位点(locus control region, LCR)的控制。
对非珠蛋白LCR的研究使我们对LCR的结构和功能有了进一步的认识。LCR由一系列组织特异性的HS位点组成。这些HS位点并非必需像β-珠蛋白LCR的HS位点那样分布在一段连续的DNA片段上。它们可以位于基因簇的上游、下游或者基因之间。每一HS位点包含一个150~300 bp的核心序列,其中有很多转录因子的结合位点。人类CD2 LCR是建立开放的染色质结构所必需的,但是没有增强子的功能。
10.1.4.4 沉默子 在酿酒酵母中,HML和HMR位点,以及接近端粒的染色质区域,形成一种抑制性染色质结构,因此在转录上是沉默的。HM位点上的沉默效应是由位点两侧、短的被称为沉默子的特异序列起始的。
沉默子结合蛋白通过募集Sir沉默复合体起始沉默过程。Sir沉默复合体由Sir2, Sir3和Sir4构成。Sir2是一种NAD-依赖型组蛋白去乙酰化酶,Sir3和Sir4为组蛋白结合蛋白。一旦被募集到沉默子,Sir复合体使附近的核小体脱乙酰化。由于Sir复合体自身的相互作用,以及Sir复合体优先结合于乙酰化程度低的组蛋白,新的Sir复合体就会被募集到刚刚脱去乙酰化的核小体上。于是,新一轮的核小体去乙酰化、Sir复合体与去乙酰化的核小体结合的过程又开始了。按照这种方式,Sir复合体介导核小体的去乙酰化作用逐渐地、沿染色质扩散,导致HM位点的异染色质化。
染色体的端粒重复序列含有一系列Rap1p的结合位点。Rap1p募集Sir复合体,起始异染色质的生成和扩散,最终在染色体的近端粒区形成沉默的染色质结构。染色体的端粒重复序列含有一系列Rap1p的结合位点。Rap1p募集Sir复合体,起始异染色质的生成和扩散,最终在染色体的近端粒区形成沉默的染色质结构。
10.2 DNA甲基化与基因组沉默 10.2.1 真核生物DNA的甲基化 DNA甲基化是指在DNA甲基化酶(DNA methyltransferase)的作用下,以S-腺苷甲硫氨酸为甲基供体,将甲基转移到DNA分子的胞嘧啶碱基上形成5-甲基胞嘧啶的过程。胞嘧啶的甲基化作用不是随机的。在脊椎动物基因组中胞嘧啶甲基化仅限于5’-CG-3’二核苷酸,植物中仅限于5’-CG-3’二核苷酸和5’-CNG-3’三核苷酸。
脊椎动物基因组中仅有不到1/4的CpG位点得以保留。脊椎动物基因组中仅有不到1/4的CpG位点得以保留。
CpG岛 在基因组中,CpG二核苷酸并非随机分布,基因组的某些区域其CpG二核苷酸的水平比平均值高10~20倍,这些CpG富集区被定义为CpG岛。
10.2.2 DNA甲基化与基因沉默 基因沉默是指细胞以相对非特异性的方式关闭基因的表达,它可以影响一个基因、一个基因簇、染色体的一个区段甚至整条染色体。 DNA甲基化可以引起基因沉默。
甲基化的生物学效应是由各种mCpG结合蛋白(methyl-CpG-binding proteins, MeCP)介导的。
10.2.3 DNA甲基化与基因组印记 在一个二倍体细胞中,常染色体基因有两个拷贝,一个来自父本,一个来自母本。多数情况下,两个基因在功能上是等价,它们在表达水平上具有可比性。但在少数情况下,二倍体细胞核中同源染色体上的一对等位基因中只有一个可以表达,另一个因甲基化而沉默,这种现象就是基因印记,就如同基因被打上了亲代的印记。对一些印记基因来说,来源于父本的等位基因不表达,来源于母本的基因表达。对另一些印记基因来说,情况刚好相反。






















