Download

1 / 33
390 likes | 858 Views
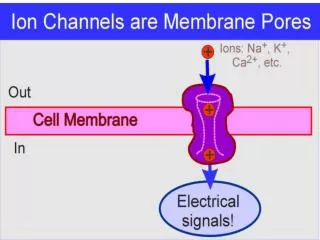
Ion Channels. Active Transporters: The proteins that created and maintain ion gradients Ion channels : give rise to selective ion permeability changes. ION CHANNELS.

E N D
Active Transporters: The proteins that created and maintain ion gradients Ion channels : give rise to selective ion permeability changes
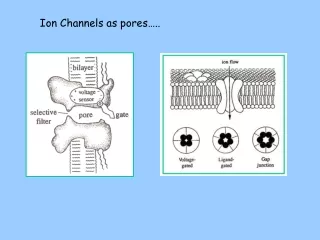
ION CHANNELS Ion channels are transmembrane proteins that contain a specialized structure, THE PORE that that allow particulars ions to cross the membrane. Some ion channels contain voltage sensor ( voltage gated channels) that open or close the channel in response to changes in voltage. Other gated channels are regulated by extracellular chemical signals such as neurotransmitter or by intracellular signals as a second messengers.
ACTIVE TRANSPORTERS Membrane proteins that produce and maintain ion concentration gradients. For example the Na+ pump which utilizes ATP to regulate internal concentration of Na+ and K+. Transporters create the ionic gradient that drive ions through open channels, thus generating electric signals
What is the mechanism for ion movement across the membrane? • K+ and Na+ currents were distinct, suggesting distinct mechanisms • Mechanism is voltage dependent (must sense voltage) • Voltage clamp recordings showed that ions move across membrane at high rates (~ 600,000 /s) – inconsistent with an ion pump mechanism • Ion selectivity of Na+ and K+ currents – size dependent permeability suggests pore of certain diameter. • Armstrong (1965-6) – TEA block could be overcome by adding excess K+ to the extracellular fluid and stepping to hyperpolarized potentials (K+ comes into cell) suggesting that K+ ions dislodge TEA from pore
Ion channels share several characteristics • The flux of ions through the channel is passive . • The kinetic properties of ion permeation are best described by the channel conductance (g) that is determinate by measuring the current flux (I) that flows through the channel in repose to a given electrochemical driving force. (Electrochemical driving force is determinate by difference in electric potential across membrane and gradient of concentration of ions) . At the single channel level, the gating transitions are stochastic. They can be predicted only in terms of probability.
Ion channels share several characteristics • In some channels the current flow varies linearly with the driving force ( channels behave as resistors) • In other channels, current flow is a non-linear function of driving force ( Rectifiers) High conductance (γ) I (pA) V (mV) Low conductance (γ) Ohmic channel Rectifying Channel ( I=Vm/R)
Ion channels share several characteristics • The rate of ion flux (current) depends on the concentration of the ions in solution ( At low concentrations the current increases linearly with the concentration, at higher concentrations the current reach a saturation point ) . • The ionic concentration at which current flow reaches half its maximum defines the dissociation constant for ion binding. • Some ion channels are susceptible to occlusion by free ions or molecules
The Opening and closing of channels involve conformational changes • In all channel so far studied, the channel protein has two or more conformational states that are relatively stable. Each stable conformation represents a different functional state.. Each channel has an open state and one or two closed states. The transition between states is calling gating.
The Opening and closing of channels involve conformational changes • Three major regulatory mechanisms have evolved to control the amount of time that a channel remains open and active. • Under the influence of these regulators ,channels enter one of three functional states: closed and activable (resting), open (active) or closed and nonactivable ( refractory). • The signal that gate the channel also controls the rate of transition between states.
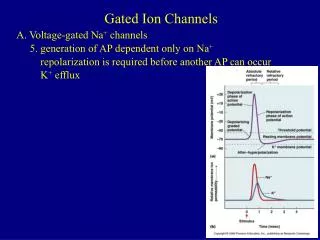
The Opening and closing of channels involve conformational changes • Ligand -gated and voltage gated channels enter refractory states through different process. Ligand-gated channels can enter refractory state when the exposure to ligand sis prolonged (desensitization) • Voltage-gate channels enter a refractory state after activation. The process is called inactivation. Activation is the rapid process that opens Na+ channels during a depolarization. Inactivation is a process that closes Na+ channels during depolarization. The membrane needs to be hyperpolarized for many milliseconds to remove inactivation.
The Opening and closing of channels involve conformational changes • Exogenous factors such as drugs and toxins can affect the gating control sites.
Structure of Ion Channels • Ion channels are composed of several subunits. They can be constructed as heterooligomers from distinct subunits, as homooligomers from a single type of subunit o from a single polypeptide chain organized into repeated motifs. • In addition to one or more pore forming unties, which comprise a central core, some channels contain auxiliary subunits which modulate the characteristics of the central core
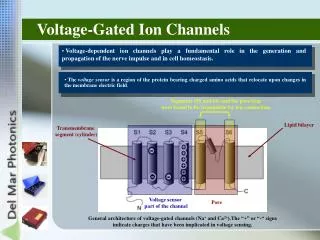
Structure of voltage gated ion channels Repeated series of 6 TM a helices S4 helix is voltage sensor Loop between S5 & S6 composes selectivity filter
Gating currents Movement of + charges in S4 segment produces small outward current that precedes ion flux through channel
Role of auxiliary subunits Auxiliary (non pore) subunits affect: • Surface expression • Gating properties
Voltage gated sodium channels A large alpha subunit that forms the core of the channel and its functional on its own. It can associate with beta subunits Blocked by: TTX, STX, *cain local anesthetics
Persistent (non-inactivating) Na+ currents are produced by an alternative channel gating mode
Voltage gated Ca2+ channels Gene Product Cav1.1-1.4 Cav2.1 Cav2.2 Cav2.3 Cav3.1-3.3 Tsien Type “L” “P/Q” “N” “R” “T” Characteristics High voltage Mod voltage High voltage Mod voltage Low voltage activated, activated, activated, activated activated slow inactivation moderate moderate fast fast (Ca2+ dependent) inactivation inactivation inactivation inactivation Blocked by dihydropiridines Agatoxin Conotoxin SNX 482 Mibefridil (nimodipine) IVA GVIA High Ni2+ Form by different subunits:α1, α2δ,β and γ. The α1 subunit forms the pore, the other subunits modulate gating.
Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ dependent Ca2+ channel inactivation Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ Ca2+ channel CaM -
Inwardly rectifying Voltage gated 2 pore (“leak”) Ca2+ activated Potassium Channels
Inwardly-rectifying 2 pore “leak” Inwardly-rectifying channels • subunits: Kir 1.X - 7.X • Rectifying character due to internal block by Mg2+ and polyamines • Roles: • Constitutively active resting K+ conductance (eg. Kir1, Kir2) • G-protein activated (Kir3) • ATP sensitive (Kir6) 2 pore “leak” channels • many different subunits, nomenclature still argued • Outwardly rectifying due to unequal [K+] across the membrane • Roles: • Constitutively active resting K+ conductance • pH sensing • Mechanosensitive • Thermosensitive • Second messenger sensitive (cAMP, PKC, arachadonic acid) Inwardly-rectifying and “leak” K+ channels
Voltage gated K+ channels Gene Product Kv1.X (1-8) Kv2.X (1-2) Kv3.X (1-4) Kv4.X (1-4) Kv7.X (1-5) “D type” “Delayed “Delayed “A type” “M current” rectifier” rectifier” Characteristics Low voltage High voltage High voltage Low voltage Low voltage activated (~50 mV), activated (0 mV), activated (-10 mV), activated (-60 mV) activated (-60 mV) fast activation mod activation fast activation fast activation slow activation (< 10 ms) (>20 ms) (10-20 ms) (10-20 ms) (>100 ms) slow inactivation very slow very slow fast no inactivation inactivation inactivation inactivation fast deactivation Blocked by 4-AP (100 µM) TEA (5-10 mM) TEA (0.1-0.5 mM) 4-AP (5 mM) XE991 dendrotoxin 4AP (1-5 mM) 4AP (0.5-1 mM) BDS (50 nM) Kv4 (“A type”) Kv1 (“D type”) Kv2 (“DR type”) Kv3 (“DR type”)
Ca2+ activated K+ channels - role in repolarization following APs Voltage response currents mediating AHP Spike frequency accommodation Role of IKCa in burst duration
Ca2+ activated K+ channels Channel Type BK SK sAHP “maxi K, IC fAHP” “mAHP” “sAHP” Gene product slo 1-3 SK1-3 ???? Voltage dep? Yes No No [Ca2+] to activate 1-10 µM 0.1-1 µM 0.1-1 µM Ca2+ binding direct to subunit calmodulin hippocalcin? Single channel 100-400 pS 5-20 pS 5-10 pS Conductance Blocked by charybdotoxin apamin TEA (> 20 mM) TEA (< 1 mM) TEA (> 20 mM)
Effect of drugs and toxins Many toxins block ion channels directly either from the outer (TTX) or inner (lidocaine) surface of the channel Other toxins change the properties of the channel without blocking it Delaying inactivation Shifting voltage dependence FUGU TTX LA
Modulation of Ion Channels Example, enhancement of Ca2+ channels in cardiac myocytes by NE
Dendritic ion channels participate in synaptic amplification and integration
Recommended Readings: Kandel. Principles of Neural Science, 4 th Edition chapter: 6 Hille. Ion Channels of Excitable Membranes. 3 ed. Edition.