Download

1 / 49
490 likes | 596 Views
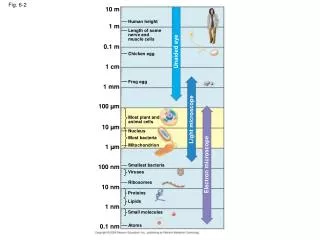
VVP Fig 23-2. MVA Fig. 28.1. MVA Fig. 28.7 A mitotic chromosome Loops are 45-95 kbp. Alberts, Figure 4-23. prolate ellipsoid 100 x 70 Angstroms. 2 turns around histone core. Nuclease experiments produce 166 bp fragments Upon loss of H1, 146 bp fragments are produced.

E N D
MVA Fig. 28.7 A mitotic chromosome Loops are 45-95 kbp
2 turns around histone core Nuclease experiments produce 166 bp fragments Upon loss of H1, 146 bp fragments are produced 1.45 turns around histone core Smaller fragments correspond to loss of H1.
Properties of histones residues MW %arg %lys H1 215 23.0 1 29 H2a 129 14 9 11 H2b 125 13.8 6 16 H3 135 15.3 13 10 H4 102 11.3 14 11
Protein component of nucleosome • 8 histone monomers • 2 each of H2A, H2B form two heterodimers • 2 each of H3, H4 form heterodimers that then tetramerize
Three “Domains” of histones • N-terminal tails • Post-translational modifications • Canonical histone fold • Segment-swapped • Four-helix bundle tetramer interface Representative archael histone: hMfB JReeve et al., Ohio State University
For each dimer: 2 unfolded monomers intermediate partiallly native dimer folded dimer It is likely that the intermediate form is what binds to the DNA and then folds the rest of the way.
14 different arg stick their side chains into the minor grove!
VVP Fig. 27-31 Chromosome Puffs where transcription is occurring.
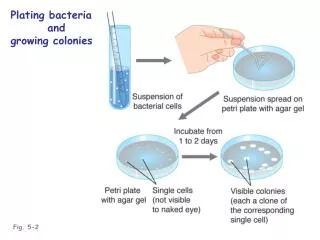
Demonstration that transcriptionally active genes are more susceptible than inactive genes to DNase I digestion. Chick embryo erythroblasts at 14 days actively synthesize globin, whereas cultured undifferentiated MSB cells do not. Nuclei from each type of cell were isolated and exposed to increasing concentrations of DNase I. The nuclear DNA was then extracted and treated with the restriction enzyme BamHI, which cleaves the DNA around the globin sequence and normally releases a 4.6-kb globin fragment (a). The DNase I- and BamHI-digested DNA was subjected to Southern-blot analysis with a probe of labeled cloned adult globin DNA, which hybridizes to the 4.6-kb BamHI fragment. As shown in (b) the transcriptionally active DNA from the 14-day globin-synthesizing cells was sensitive to DNase I digestion, indicated by the absence of the 4.6-kb band at higher nuclease concentrations. In contrast, the inactive DNA from MSB cells was resistant to digestion. [See J. Stalder et al., 1980, Cell 19:973.]
Experimental method for analyzing the acetylation state of histones in chromatin associated with a specific region of the genome. Nucleosomes are lightly cross-linked to DNA in vivo using a cell-permeable, reversible, chemical cross-linking agent. Chromatin is then isolated, sheared to an average length of three nucleosomes, and subjected to immuno-precipitation with an antibody specific for a particular acetylated N-terminal histone sequence. The DNA in the immunoprecipitated chromatin fragment is released by reversing the cross-link and then is quantitated using a sensitive PCR method. [See S. E. Rundlett et al., 1998, Nature 392:831.]
(H3H4 )2 less stable than H2A-H2B H2A-H2B depleted in actively transcribing chromatin. H3H4 stays with the DNA once positioned. Since it’s less stable, polymers are freer to get out of the way of the transcription apparatus
Bromodomain bound to Acetyl-lys of H4 These 110 aa domains are presentin many proteins that regulate eukaryotic transcription.
Chromatin structure and gene expression. Organization of chromatin into the tightly condensed 30-nm fibre and 'beads-on-a-string' 10-nm fibre. b | Region of DNA containing an actively transcribed gene. The region encompassing the transcribed gene probably adopts the 10-nm configuration, whereas sequences up- and downstream are organized into a 30-nm fibre. Transcription involves changes in the chromatin structure that can be detected by digestion with the endonuclease DNaseI. The transcribed regions also contain hypersensitive (HS) sites, which may be associated with a small stretch of DNA devoid of nucleosomes or simply reflect a different nucleosomal structure (positioning or histone modification). They usually correlate with non-histone proteins bound to the DNA. Regions of general DNaseI sensitivity correlate with acetylation of the histone tails, defining a structural and functional 'domain' for gene activity that is characteristic of 'open' chromatin. This domain is embedded in regions of higher chromatin compaction that are more resistant to DNaseI digestion. This compacted structure correlates with the presence of deacetylated histones (blue wavy lines), and is characteristic of 'closed' chromatin.
Models for the generation and propagation of histone codes. Recruitment of chromatin modifiers including histone acetyl transferases (HATs), histone deacetylases (HDACs) and histone methyl transferases (HMTs) (coloured boxes) through sequence-specific DNA-binding factors (DBFs; coloured ovals) generate chromatin codes in the vicinity of their cognate sites (similarly colored rectangles on the DNA ribbon). Histone modifiers that generate codes consistent with gene inactivity are shown in orange, whereas those that correlate with gene activity are shown in green.
Histone modifications as chromatin codes. Schematic representation of histone modifications encountered on inactive and active gene loci. A. Acetylation (Ac) of lysine (K) 12 on histone H4 and methylation (Me) of K9 on H3 are found at inactive gene loci and are also enriched in heterochromatin (orange circles and squares). B. Acetylation on K9 and K14 of H3, methylation on K4 of H3, acetylation on K5 of H4, and methylation on arginine 3 of H4 are found at gene loci that are either active or have the potential to be (green circles and squares).