Download

1 / 25
250 likes | 394 Views
Supply-demand for NADPH. Celton at al. (2012) A constraint-based model analysis of the metabolic consequences of increased NADPH oxidation in Saccharomyces cerevisiae . Metab Eng 14:366-79

E N D
Supply-demand for NADPH Celton at al. (2012) A constraint-based model analysis of the metabolic consequences of increased NADPH oxidation in Saccharomyces cerevisiae. Metab Eng 14:366-79 Celton et al. (2012) A comparative transcriptomic, fluxomic and metabolomic analysis of the response of Saccharomyces cerevisiae to increases in NADPH oxidation. BMC Genomics 13:317
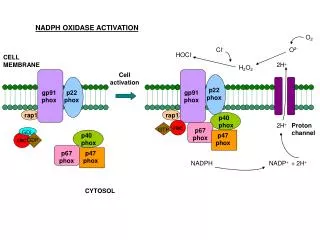
Cosa succede se si aumenta il demand di NADPH? Principali vie che producono NADPH in lievito
Spingendo la produzione di glicerolo si cercava di ridurre la resa in etanolo Engineered Saccharomyces cerevisiae strains overexpressing glycerol-3-phosphate dehydrogenase, and lacking the acetaldehyde dehydrogenase Ald6 display large-scale diversion of the carbon flux from ethanol toward glycerol without accumulating acetate Ehsani et al. (2009) Engineering of 2,3-butanedioldehydrogenase to reduce acetoin formation by glycerol over-producing, low-alcohol Saccharomyces cerevisiae. Appl.Environ.Microbiol.75, 3196–3205. purtroppo si produce Acetoino che conferisce un gusto sgradevole GlyP Acetoin is reduced to 2,3-butanediol by the NADH-dependent 2,3-butanediol dehydrogenase Bdh1 Bdh1 overexpression
Intracellular redox potential is determined principally by the ratio of NADH/NAD+ and NADPH/NADP+ cofactors, which are involved in about 200 reactions in Saccharomyces cerevisiae. These metabolites are interconnected by many different pathways. The anabolic demand for NADPH is coupled to the rate of biomass formation, which can vary significantly with environmental conditions Different levels of NADPH demand were obtained by expressing a specific dehydrogenase coupled to the addition increasing quantities of the relative substrate (acetoin) to the medium. Microarray analysis of NADPH-Bdh cells with various levels of NADPH demand We cultured the NADH-Bdh strain in the presence of 200 mM acetoin, both as a control and to assess the specificity of the effect on NADPH demand. Use of a constraint-based approach based on DynamoYeast, a dedicated stoichiometric model of S. cerevisiae during fermentation
Supply Demand NADP+ NADPH Bdh1p Increase NADPH demand by adding acetoin to the growth medium glycerol-DHA cycle Overexpressing an engineered NADPH-dependent Bdh1p enzyme in the presence of acetoin Which patway(s) for the NADPH supply?
BDH specific activity Growth of strains under oxidative stress Strain NADPH-Bdh has increased sensitivity to oxidative stress, suggesting that over-expression of NADPH-Bdh enhances the in vivo NADPH oxidation. (A) BDH specific activity of strains 59A (control) and 59A overexpressing the native (NADH-Bdh) or modified (NADPH-Bdh) BDH, determined in the presence of 20mM of NADH (black) or NADPH (gray) and (B) growth of 59A, NADH-Bdh and NADPH-Bdh strains under oxidative stress conditions. Serial dilutions of yeast cultures were spotted onto SD medium containing 100mM acetoin and 1-3mM H2O2
The Bdh-overproducing strains consumed 2–3-fold more acetoin than the control strain 59A, regardless of the initial acetoin concentration NADH-Bdh NADPH-Bdh 59A Tutto l’acetoino è consumato Acetoin and NAD(P)H consumed (mM) by the NAD(P)H-Bdh system as a function of acetoin added, for strains 59A(red), NADH-Bdh (green) and NADPH-Bdh (blue),
These conditions increase NADPH demand by factors of 8, 13 and 22, respectively, with respect to the anabolic demand. Cosa succede al resto del metabolismo? Quantification of the NADPH perturbations Comparison between the amount of NADPH consumed (normalized to biomass, mmol/gDW) by the anabolic demand, determined from biomass composition (yellow), and via the NADPH-Bdh system (dark blue)
Con l’enzima NADH dipendente, quanto aumenta il consumo di NADH? E quanto il consumo per la sintesi di glicerolo nel wt? glycerol pathway in wt NADH-Bdh system Quantification of the NADH perturbations Comparison between the amount of NADH oxidized (normalized to biomass, mmol/gDW) by the glycerol pathway (light blue) and by the NADH-Bdh system (green).
increased excretion of acetate and acetaldehyde The greater acetate formation may there- fore allow compensation, at least in part, for the consumption of NADPH La causa della produzione di acetato sono diverse nei due ceppi BDH
acetate (C) Growth (A) NADH-Bdh NADPH-Bdh NADH-Bdh NADPH-Bdh 0 mM 400 mM 400 mM 0 mM glycerol (B) succinate (D) The response to NADH perturbation was a gradual decrease in glycerol formation, consistent with the role of glycerol in maintaining NADH homeostasis Presence of 0 (green), 100 (turquoise), 200 (purple), 300 (orange) and 400 mM (magenta) acetoin
Also a trancriptome analysis on cells harvested at mid-exponential growth phase when the amount of cumulative CO2 release was 6 g/l. 200 (NADH-Bdhp open triangle) without acetoin (59A, circle) 0 (circle) 100 (square) 200 (triangle) (NADPH-Bdhp) Growth 300 (diamond) mM acetoin Cells sampled when CO2 reached 6 g/L 0 (circle) All the strains grew at the same rate, except NADPH-Bdh in the presence of 300 mM acetoin, which had a growth rate decreased by 60%
At this time point, strains exposed to different degrees of change in NADPH demand differed in their acetate (a marker of NADPH metabolism) production 300 (diam) 200 (triangle) 100 (square) 200 NADH-BDH (open triangle) 0 (circle) Acetate production mM acetoin Cells were sampled when CO2 reached 6 g/L E’ possibile aumentare la richiesta di NADPH di molte volte rispetto al livello basale con piccole alterazioni nel metabolismo aumenta la produzione di acetato
Complete sugar exhaustion was achieved in 36 h for 59A, in 36, 46 and 54 h for NADH-Bdhp in the presence of 100, 200 and 300 mM acetoin, and in 50 h for NADH-Bdhp in the presence of 200 mM acetoin. NADPH anabolic demand can be increased 8 times without any ill effect on growth The pentose phosphate pathway (PP pathway) and the acetate synthesis pathway (via the action of the NADP+-dependent acetaldehyde dehydrogenase Ald6p) satisfied 80 and 20%, respectively, of the NADPH demand when this demand was increased to up to 22 times the anabolic requirement. If demand was increased still further (40 times the anabolic demand), the PP pathway was saturated and our model predicted a role for the glycerol-DHA cycle, which exchanges NADP+ and NADH for NAD+ and NADPH, at the expense of one ATP molecule.
Our findings reveal that S. cerevisiae responds robustly to perturbations of between 8 and 40-fold in the NADPH demand for anabolism. The predictions of the model highlight three points: (1) The contribution of mitochondrial shuttles resulting in the net production of NADH in the cytosol, explaining the production of a glycerol surplus with respect to the anabolic demand; (2) The major contributions of the PP and, to a lesser extent, acetate pathways to providing NADPH in response to increased NADPH demand (3) The saturation of the PP pathway at high levels of NADPH oxidation, leading to reducing equivalents being transferred from NADH to NADPH at the expense of ATP by means of the glycerol–DHA cycle.
Despite these significant advances in our understanding of NADPH metabolism, little is known about the mechanisms regulating NADPH homeostasis. It is generally thought that the pentose phosphate pathway is controlled principally at the enzymatic level, with NADPH and ATP competitively inhibiting both the glucose-6 phosphate dehydrogenase Zwf1p and the 6-phosphogluconate dehydrogenase Gnd1p. The coordinated regulation of genes involved in NADPH metabolism, including most of PP pathway genes, has been reported in conditions of oxidative stress. The activation of NADPH-dependent genes involves Stb5p, a zinc-binding factor, which also represses the expression of PGI1, encoding the phosphoglucose isomerase at the junction between glycolysis and the PP pathway. This transcription factor plays a key role in rerouting carbon flux to provide the additional NADPH required for the response to oxidative stress, as demonstrated by the greater susceptibility of the stb5Δ mutant to several chemicals and oxidants and by the greater resistance to diamide of a strain overexpressing STB5. Stb5p was also recently shown to be essential for acetaldehyde tolerance and, under anaerobic conditions, to play a major role in the maintenance of basal flux through the PP pathway.
The number of genes displaying differential expression clearly increased with the degree to which NADPH demand was modified. In the presence of 100 mM acetoin, only eight genes displayed a change in expression level, whereas 71 genes were upregulated and 37 were downregulated in the presence of 200 mM acetoin and 61 genes were upregulated and 48 were downregulated in the presence of 300 mM acetoin. Changes in NADH demand had little overall effect on gene expression, as only 23 genes were differentially regulated. This suggests that the response to an increase in NADH demand to four times (at 200 mM acetoin) or in NADPH to eight times (at 100 mM acetoin) the amount required for anabolism mostly involved metabolic control. Upregulated Downregulated
Transcriptomic response of PP pathway genes to increases in NADPH demand. Induction levels (fold-changes) Upregulated genes in red metabolic control
Transcriptomic response of genes of the sulphur assimilation pathway in various NADPH demand conditions Sulfate assimilation requires considerable amounts of NADPH (7 moles of NADPH are oxidized to NADP+ to generate 1 mole of methionine from 1 mole of SO42−).
Figure 6 Intracellular metabolite concentrations for the glycolysis, PP and carbohydrate pathways
Dall’analisi dei flussi, dei dati di trascrittomica e delle concentrazioni dei metaboliti, l’aumento del consumo di NADPH di 8 volte rispetto al consumo basale non ha quasi alcun effetto sul metabolismo. All’aumento di richiesta di NADPH, il sistema risponde "automaticamente" aumentando il flusso. Control by demand