Download

1 / 42

E N D
Chemiosmotic Theory M.Prasad Naidu MSc Medical Biochemistry, Ph.D,.
Theories of oxidative phosphorylation • Chemiosmotic theory • Boyer’s binding change mechanism
The Chemiosmotic Theory of oxidative phosphorylation, for which Peter Mitchell received the Nobel prize: Coupling of ATP synthesis to respiration is indirect, via a H+ electrochemical gradient.
Chemiosmotic theoryproposed byPeter Mitchell The transport of protons from matrix to intermembrane space is accompanied by the generation of a proton gradient across the membrane.
Protons (H+) accumulate intermembrane space creating an electrochemical potential difference, proton gradient or electrochemical gradient. • This proton motive force (PMF) drives the synthesis of ATP by ATP synthase complex.
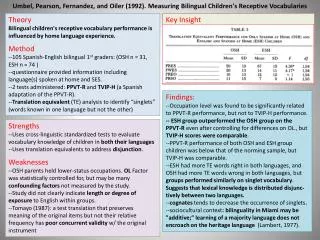
CHEMIOSMOTIC THEORY Peter mitchel IMM- Inner mitochondrial membrane IMS- Inter membrane space OMM- outer mitochondrial membrane H+ H+ H+ H+ 4H+ H+ 4H+ H+ 2H+ III 2e- H+ 2e- H+ I H+ Iv 4H+ H+ H+ H+ H+ 2H+ 4H+ H+ H+ H+ ADP+Pi H+ 4H+ H+ H+ 4H+ H+ V H+ H+ H+ H+ ATP H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ MATRIX H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ IMM H+ H+ H+ H+ H+ Complex I, III and IV are proton pumps H+ H+ IMS H+ OMM
Proton gradient / electrochemical gradient • Proton motive force
Generation of ATP • Proton dependant ATP synthese • Uses proton gradient to make ATP • Protons pumped through channel on enzyme • From intermembrane space into matrix • ~4 H+ / ATP • Called chemiosmotic theory
Totals NADH 10 H+ X 1 ATP = 2.5 (3) ATP 4 H+ FADH2 6 H+ X 1 ATP = 1.5 (2) ATP 4 H+
Boyer ’s binding change mechanism: ATP synthase is a protein assembly in the inner mitochondrial membrane.
ATP synthase has two units F1 - projects into matrix -has 3 α , 3 β , gamma , delta, epsilon chains -catalyses ATP synthesis Peripheral catalytic sites are present on beta subunits. Fo - embedded in membrane - acts as channel for transport of H+
4 H+ H+ H+ H+ H+ H+ H+ H+
Mechanism of ATP synthesis (Boyer’s Hypothesis) • Boyer’s binding change hypothesis • Synthesis of ATP occurs on the surface of F1. • Binding change mechanism states that 3 beta • subunits change CONFORMATIONS during • catalysis with only one beta subunit acting as • Catalytic site.
β subunits occur in 3 forms • ‘O’ form (Open form). It has low affinity for substrates ADP +Pi • ‘L’ form (loose form). Can bind substrates ADP and Pi but catalytically it is inactive. • ‘T’ form (Tight form). Binds substrates ADP + Pi tightly and catalyses ATP synthesis.
When protons pass through the disk of C subunits of F0 unit it causes rotation of γ sub unit. • The β subunits which are fixed to the membrane donot rotate. • ADP & Pi are taken up sequentially by the βsubunits which undergo conformational changes and form ATP.
Gamma subunit is in the form of axle . It rotates when protons flow. ATP synthase is smallest known MOLECULAR MOTOR in the living cells.
ETC - inhibitors Complex I : site I of ATP synthesis inhibitors Rotenone, Peircidin, Amytal, Barbiturates ComplexII: Carboxin,Thenoyltrifluroacetone,malonate Complex III : site II of ATP synthesis inhibitors Antimycin, Myxothiazol , stigmatellin Complex IV: site III of ATP synthesis inhibitors Cyanide, azide , carbon monoxide
Complex – I inhibitors (Site I inhibitors) Rotenone, insecticide, also used as fish poison. Binds to complex I and prevents the reduction of Ubiquinone. Piercidin, Amytal (sedative), Barbiturates – inhibit by preventing the transfer of electrons from iron sulfur center of complex – I to Ubiquinone.
Complex – II inhibitors Malonate acts as a competitive inhibitor with the substrate succinate
Complex – III inhibitors (Site II Inhibitors) Antimycin inhibit electron transfer from cytb to C1. Myxothiazol and stigmatellin, antibiotics inhibit electron transfer from Cytb to C1.
Complex – IV (site III inhibitors) Cyanide and azide bind tightly to oxidized form of heme a3 ( of complex iv ) preventing electron flow. Cyanide is potent and rapidly acting poison. Cyanide prevents binding of oxygen to Cytochrome oxidase ( aa3). Mitochondrial respiration and energy production stops cell death occurs rapidly.
Carbonmonoxide binds to the reduced form of heme a3(Fe2+) competitively with oxygen and prevents electron transfer to oxygen.
Uncouplers of oxidative phosphorylation : • Uncouplers will allow oxidation to proceed but energy instead of being trapped as ATP is dissipated as heat. • They are hydrophobic weak acids. • They are protonated in the intermembrane space where a higher concentration of protons exists.
These protonated uncouplers due to their lipophilic nature rapidly diffuse across the membrane into matrix where they are deprotonated since matrix has a lower concentration of protons. • Thus, the proton gradient is dissipated.
2-4 dinitrophenol a classical uncoupler – electrons from NADH to oxygen proceeds normally but ATP not formed as proton motive force across inner mitochondrial membrane is dissipated .
2. Penta chloro phenol 3. Dinitro cresol 4.Bilirubin 5.Thyroxine-Physiological uncoupler 6.Valinomycin 7.Nigericin Note: They are Lipophilic
Intermembrane space matrix H+ H+ H+ H+ H+ H+ H+ H+ H+ H+
Physiological Uncouplers 1.Excessive thyroid hormones 2. Unconjugated hyper bilirubinaemia 3. In high doses aspirin uncouple oxidative phospharylation which explains fever that accompanies toxic over dosage of these drugs.
Uncoupling proteins • UCPs occur in the inner mitochondrial membrane of mammals, including humans. • UCPs create a “proton leak”, that is they allow protons to re-enter the mitochondrial matrix without energy being captured as ATP. • Energy is released as heat, and the process is called nonshivering thermogenesis.
UCP1, also called thermogenin, is responsible for the activation of fatty acid oxidation and heat production in the brown adipocytes of mammals. • Brown fat , unlike the more abundant white fat, uses almost 90% of its respiratory energy for thermogenesis in response to cold, at birth,etc.
Inhibitors of Oxidative phosphorylation : • Oligomycin – acts through one of the proteins present in F0 - F1 stalk . • Oligomycin blocks the synthesis of ATP by preventing the movement of protons through ATP synthase.
Respiratory control The regulation of the rate of oxidative phosphorylation by ADP level is called respiratory control. The ADP level increases when ATP is consumed and so oxidation is coupled to the utilization of ATP. Under physiological conditions, electron transport is tightly coupled to oxidative phosphorylation.
Electrons do not usually flow through the electron transport chain to O2 unless ADP is simultaneously phosphorylated to ATP. In the presence of excess substrate and Oxygen, respiration continues until all ADP is converted to ATP. After that the respiration rate or utilization of oxygen decreases In the presence of adequate oxygen and substrate, ADP becomes rate limiting; it exerts a control over the entire oxidative phosphorylation process
The rate of respiration of mitochondria (Oxidative phosphorylation) can be controlled by ADP. • Oxidation cannot proceed via ETC without simultaneous phosphorylation of ADP. • Chance & Williams defined 5 conditions that can control rate of respiration.
Generally most cells in the resting state are in state 4 , and respiration is controlled by the availability of ADP. • The availability of inorganic phosphate could also influence the respiration. • As respiration increases (Exercise) cell approaches state 3 ( ETC working to its full capacity ) or state 5 ( Availability of O2 is a limiting factor ). • ADP / ATP transporter may also be a rate limiting factor
P:O ratio (ADP : O ratio) P:O ratio is defined as number of phosphates incorporated into ATP to 1 atom of oxygen utilized during the transfer of 2 electrons through ETC. For NADH P:O ratio is 3 i.e 3 ATPs are produced (2.5) For FADH2 P:O ratio is 2 i.e 2 ATPs are produced(1.5)