Download
1 / 26
260 likes | 479 Views

E N D
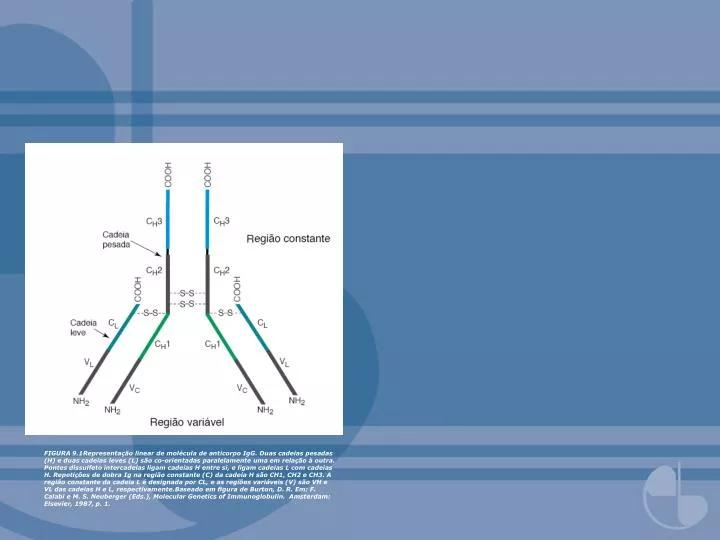
FIGURA 9.1Representação linear de molécula de anticorpo IgG. Duas cadeias pesadas (H) e duas cadeias leves (L) são co-orientadas paralelamente uma em relação à outra. Pontes dissulfeto intercadeias ligam cadeias H entre si, e ligam cadeias L com cadeias H. Repetições de dobra Ig na região constante (C) da cadeia H são CH1, CH2 e CH3. A região constante da cadeia L é designada por CL, e as regiões variáveis (V) são VH e VL das cadeias H e L, respectivamente.Baseado em figura de Burton, D. R. Em: F. Calabi e M. S. Neuberger (Eds.), Molecular Genetics of Immunoglobulin. Amsterdam: Elsevier, 1987, p. 1.
FIGURA 9.2Estrutura diagramática de IgG. Cadeias leves (L) são divididas em dobras Ig VL (seqüência variável de aminoácidos) e CL (seqüência constante de aminoáci-dos). Cadeias pesadas (H) são divididas em domínios VH e CH1, CH2 e CH3. Sítios de liga-ção de antígeno são VH-VL. Polipeptídeos de articulação (hinge) interconectam do-mínios. Posições de ligações cistina inter- e intra-cadeia são mostradas.De Cantor, C. R. e Schimmel, P. R. Biophy-sical Chemistry. Parte I. San Francisco: Freeman, 1980. Reimpresso com permissão de Sr. Irving Geis, New York.
FIGURA 9.4Dobra de Imunoglobulina. (a) Diagra-ma esquemático de dobramento de um domínio CL, mostrando a estrutura em folha-β pregueada. Setas indicam fitas de folha-β, e barra (cinza-claro) indica posição de ligação cistina. Setas cinza são fitas-β no plano acima, e setas pretas são fitas-β no plano abaixo. (b) Esquema diagramático do arranjo (topologia) de fitas-β no motivo de dobra de imunoglobulina. Exemplos são as regiões variável (diagrama superior) e constante (diagrama inferior) de IgG. Setas grossas indicam fitas-β e linhas finas, alças que conectam fitas-β. Círcu-los indicam cisteínas que formam ponte dissulfeto intradomínio. Quadrados mostram posições de resíduos de tripto-fano que são componentes invariáveis do núcleo da dobra de imunoglobulina. Letras em negrito indicam fitas que formam um plano da folha, enquanto outras letras formam o plano paralelo, atrás do primeiro plano.Parte (a) reproduzido com permissão de Edmundson, A. B., Ely, K. R., Abola, E. E., Schiffer, M. e Pavagiotopoulos, N. Biochemistry 14:3953, 1975. Direitos autorais (1975) American Chemical So-ciety. Parte (b) redesenhado de Calabi, F. Em: F. Calabi e M. S. Neuberger (Eds.), Molecular Genetics of Immunoglobulin. Amersham: Elsevier, 1987, p. 203.
FIGURA 9.5Estrutura do carbono () do fragmento Fab de IgG KOL, mostrando domínios VL-VH e CL-CH1 interconec-tados por regiões de articulação (hinge).Redesenhado com permissão de Huber, R., Deisenho-fer, J., Coleman, P. M., Matsushima, M. e Palm, W. Em The Immune System, F. Melchers e K. Rajwsky (Eds.), 27th Mosbach Colloquium. Berlim: Springer-Verlag, 1976, p. 26. FIGURA 9.6Modelo de uma molécula de anticorpo IgG. Somente os carbonos α da estrutura aparecem. As duas cadeias L são representadas por esferas cinza-claras e as cadeias H, por esferas cinza-claras. Carboi-dratos ligados aos dois domínios CH2 são coloridos em cinza e cinza-médio. As regiões CDR dos domínios VH-VL são quase pretas, nas cadeias H, e cinza-escuras, nas cadeias L. A ponte dissulfeto intercadeia, entre as cadeias L e H, é representada em bola-bastão magenta (parcialmente escondida). O heptapeptídeo de articulação entre os domínios CH1 e CH2, conectan-do as unidades Fab e Fc, é preto. O centro do sítio do complemento C1q no domínio CH2 é cinza bem claro, os sítios de aportamento de proteína A na junção de CH2 e CH3 são cinza-escuros, e o sítio de ligação de tuftsina em CH2 é cinza-médio. Tuftsina é um tetra-peptídeo natural que induz fagocitose por macrófa-gos e pode ser transportado ligado a uma imunoglo-bulina. Proteína A é uma proteína bacteriana com uma alta afinidade por imunoglobulinas.Fotografia gentilmente cedida por Dr. Allen B. Edmun-dson, de Guddat, L. W., Shan, L., Fan, Z. C. et al. FASEB J. 9:101, 1995.
FIGURA 9.7Alças hipervariáveis em imunoglobulinas. (a) Diagrama esquemático mostrando alças hipervariáveis (CDRs) em domínio VL-VH que forma o sítio de ligação ao antígeno. (b) Um corte através de um sítio de ligação ao antígeno, mostrando contribuição de diferentes CDRs, usando modelos CPK de preenchimento de espaço dos átomos. Números referem-se a números de resíduos de aminoácidos na seqüência das regiões de alça hipervariável de cadeias leves (L) e pesadas (H).Parte (a) redesenhada de Branen, C. e Tooze, J. Introduction to Protein Structure. New York: Garland Publishing, 1991, p. 187. Parte (b) redesenhada de Branden, C. e Tooze, J. Introduction to Protein Structure. New York: Garland Publishing, 1991, p. 189. Atribuído a Chothia, C. e Lesk, A. J. Mol. Biol. 196:914, 1987.
FIGURA 9.8Hidrólise de IgG em dois fragmentos Fab e um Fc por papaína, uma enzima proteolítica
FIGURA 9.10Representação diagramática da estrutura em dobra de imunoglobulinas encontradas em diferentes proteínas da superfamília de genes imunoglobulina. Proteínas apresentadas incluem cadeias pesada e leve de imunoglobulinas, receptores de células-T, proteínas da Classe I e da Classe II do complexo principal de histocompatibilidade (MHC), proteínas acessórias de células-T envolvidas em reconhecimento de MHC da Classe I (CD8) e da Classe II (CD4) e possível formação de canais iônicos, um receptor responsável pelo transporte de certas classes de imunoglobulinas através de membranas das mucosas (poli-Ig), β2-microglobulina, que se associa com moléculas da classe I, uma proteína plasmática humana com função desconhecida (α1/β-glicoproteína), duas moléculas de função desconhecida com uma distribuição tissular que inclui linfócitos e neurônios (Thy-1, Ox-2) e duas moléculas cérebro-específicas, molécula de adesão neuronal (N-CAM) e proteína 3 neurocitoplasmática (NCP3).Redesenhado de Hunkapiller, T. e Hood, L. Nature 323:15, 1986
FIGURA 9.12Diagrama esquemático de ligação de um substrato polipeptídico com sítio de ligação em uma enzima proteolítica. P5-P3’ são resíduos de aminoácidos no substrato que se ligam aos subsítios S5-S3’ na enzima, com hidrólise peptídica ocorrendo entre P1-P1’ (seta). Direção NH2-terminal da cadeia polipeptídica do substrato é indicada por N, e direção COOH-terminal por C.Redesenhado de Polgar, L. Em: A. Neuberger e Brockleburst (Eds.), Hydrolytic Enzymes. Amsterdam, Elsevier, 1987, p. 174. FIGURA 9.14Estrutura esquemática de domínios da proteína C, mostrando estrutura multidomínios. “GLA” refere-se aos resíduos de γ-carboxiglutâmico (indicado por estruturas em árvores), no domínio NH2-terminal, pontes dissulfeto estão indicadas por barras grossas, EGF indica posições de domínios semelhantes ao fator de crescimento epidérmico, e CHO indica posições onde resíduos de açúcar estão ligados à cadeia polipeptídica. Sítios de clivagem proteolítica que levam à ativação catalítica são mostrados por flexas. Seqüência de aminoácidos é numerada a partir da extremidade NH2-terminal e serina, histidina e aspartato do sítio catalítico são mostrados no domínio catalítico por abreviaturas de uma letra dentro de círculos, S, H e D, respectivamente.Redesenhado de Long, G. L. J. Cell Biochem. 33:185, 1987.
FIGURA 9.16Organização de éxons e íntrons em genes de serino proteases. t-PA é um ativador de plasminogênio tecidual, e NGF é um fator de crescimento neural que contém uma estrutura homóloga à de uma serino protease. Éxons estão mostrados por caixas e íntrons por linhas que fazem conexão. Posição dos códons de serina, histidina e aspartato do sitio ativo são designados por S, H e D, respectivamente. Caixas pretas, à esquerda, mostram regiões que codificam peptídeo sinal NH2-terminal, que é clivado antes da secreção. Caixas claras, à direita, representam parte da seqüência transcrita em RNA mensageiro (mRNA), mas não traduzida em proteína. Setas indicam códons para resíduos nos quais ativação proteolítica do zimogênio ocorre.Redesenhado e modificado de Irwin, D. M., Roberts, K. A. e MacGillivray, R. T. J. Mol. Biol. 200:31, 1988.
FIGURA 9.17Duas imagens da estrutura da tripsina, mostrando a estrutura dobrada em seus domínios catalíticos. Serina, histidina e aspartato do sítio ativo estão indicados em preto FIGURA 9.21Estrutura secundária e terciária características de globinas da hemoglobina. Cadeias laterais de His F8 proximal, His E7 distal e Val E11 são mostradas. Outros aminoácidos da cadeia polipeptídica são representados por posições do carbono-α apenas; as letras M, V e P referem-se às cadeias laterais metil, vinil e propionato do heme.Reimpresso com permissão de Perutz, M. Br. Med. Bull. 32:195, 1976
FIGURA 9.22Comparação da conformação de (a) mioglobina e (b) cadeia de HbA1. Estruturas gerais são muito semelhantes, exceto nas extremidades NH2-terminal e COOH-terminal.Reimpresso com permissão de Fersht, A. Enzyme Structure and Mechanism. San Francisco: Freeman, 1977, pp. 12, 13.
FIGURA 9.26Estrutura quaternária de hemoglobina mostrando interações de dobra FG e hélice C na interface 1-2. (a) Contatos de interface α1-β2 entre dobras FG e hélice C são mostrados. (b) Representação cilíndrica das subunidades α1 e β2 na hemoglobina, mostrando contatos de interface α1 e β2 entre dobra FG e hélice C, vistos de lados opostos do plano x-y de (a).Parte (a) reimpressa com permissão de Dickerson, R. E. e Geis, I. The Structure and Action of Proteins. Menlo Park, CA: Benjamim. Inc., 1969, p. 56. Parte (b) reimpressa com permissão de Baldwin, J. e Chothia, C. J. Mol. Biol. 129:175, 1979.
FIGURA 9.27Diagramas de barras e preenchimento de espaço desenhados por computação gráfica mostrando movimentos de resíduos na vizinhança do heme na transição de desoxi-hemoglobina para oxi-hemoglobina. (a) Linha preta delineia posição da cadeia polipeptídica e His F8 em hemoglobina monóxido de carbono, um modelo de oxi-hemoglobina. Linha cinza delineia posições das mesmas regiões para desoxi-hemoglobina. Posição do átomo de ferro mostrada por círculo. Movimentos são para uma subunidade α. (b) Movimentos semelhantes de posição de resíduos de aminoácidos em uma subunidade β usando diagrama de preenchimento de espaço são mostrados. Rótulos dos resíduos centrados em densidade para a conformação desoxi.Redesenhado com permissão de Baldwin, J. e Chothia, C. J. Mol. Biol. 129:175, 1979.
FIGURA 9.29Par iônico entre os grupos de cadeia lateral imidazol da His-146 e carboxilato do Asp 94 na conformação desoxi (T) de hemoglobina. Uma estrutura parcial da cadeia β mostrando o esqueleto da cadeia polipeptídica para aminoácidos 87 a 95 e 142 a 146 da cadeia β na conformação desoxi. Só as cadeias laterais de Asp 94, His 146 e da His F8 93 ligadas ao heme da subunidade β são mostradas. Átomos de oxigênio estão em cinza-escuros, átomos de nitrogênio são cinza-claros e átomos de carbono e hidrogênio são pretos. A ponte de hidrogênio (linha tracejada) é mostrada entre o N-H carregado positivamente do imidazol da βHis 146 e o oxigênio carregado negativamente do carboxilato do βAsp 94.Desenho feito com o Swiss-PdbViewer usando a estrutura PDB 1A3N
FIGURA 9.35Sítio de ligação de 2,3-bisfosfoglicerato na interface - de desoxi-hemoglobina. São mostradas as cadeias laterais carregadas positivamente de duas amônias amino-terminais das duas βVal-1, imidazólio da βHis-2, ε-amino da βLys-82 e imidazólio da βHis-143. O 2,3-bisfosfoglicerato carregado negativamente se liga ao centro do anel de grupos carregados positivamente.Reimpresso com permissão de Dickerson, R. E. e Geis, I. Hemoglobin: Structure, Function, Evolution, and Pathology. Menlo Park, CA: Benjamin-Cummings, 1983. Ilustração de Irving Geis, Geis Archives Trust. Direitos pertencem a Howard Hughes Medical Institute. FIGURA 9.37Ligação e liberação de NO por hemoglobina durante o ciclo respiratório. O modelo mostra a ligação e a dissociação de NO, O2 e CO2 enquanto uma molécula de hemoglobina faz dois ciclos completos na circulação. O primeiro ciclo envolve intermediários 1-4, e o segundo ciclo, intermediários 5-8. As conformações T e R são mostradas e os grupos SH são da cadeia lateral de βCys93. O NO é ligado diretamente a um ferro-heme ou ao SH da βCys93. As etapas-chaves no transporte de NO são (i) sua ligação inicial a um heme no intermediário 3 e transferência do heme de uma subunidade β para βCys93 no intermediário 6 (conformação R) e (ii) sua transferência para uma molécula tiol pequena X-SH no intermediário 7 (conformação T), quando hemoglobina é convertida de R para T. A molécula de hemoglobina representada pode ser apenas 1 em 1.000 moléculas de hemoglobina circulantes, devido à relativamente baixa concentração molar de NO no sangue.Redesenhado de Gross, S. S. e Lane, P. Proc. Natl. Acad. Sci. USA 96:9967, 1999.
FIGURA 9.38Estrutura de Cys93 e de Cys93-S-NO nas conformações T e R. Em todas as estruturas, a borda do heme (H, cinza-escuro) aponta para o leitor e mostra estar ligada ao imidazol da histidina proximal (F8). Os dois carbonos (cinza-médios) e enxofre (cinza) da cadeia lateral da Cys 93 são apre-sentados como modelos de preenchimento espacial. Nos painéis C e D, NO é ligado ao enxofre de βCys93 com o átomo de N colorido em cinza, e o átomo de oxigênio em preto. (A) A conformação desoxi (T) com a cadeia lateral da βCys93 (-CH2-SH) está na superfície da molécula, longe do heme. A cadeia lateral da cisteína é impedida de entrar no sítio de ligação do heme pelo par iônico, ligado por pontes de hidrogênio, formado pelo imi-dazólio da βHis146 e o carboxila-to do βAsp94 (ponte de H entre grupos mostrada em cinza-claro) no lado direito superior. (B) A conformação oxi(R) com a cadeia lateral da βCys93 apontando para o heme, longe do solvente no lado externo da molécula. A ponte salina entre βHis146 e βAsp94 é quebrada na conformação R, permitindo a dobra de βCys-SH em direção ao bolsão do heme. (C) Modelo de βCys93-SNO na conformação desoxi (T). SNO é posicionado no lado de fora, acessível para reagir com moléculas pequenas X-SH no solvente. Como em (A), a cadeia lateral da cisteína é impedida de entrar no sítio do heme pelo par iônico βHis146-βAsp94. (D) Modelo de βCys93-SNO na conformação oxi(R). SNO fica mergulhado próximo do heme e longe do solvente do lado externo. Esta confor-mação facilita a transferência de NO do ferro-heme para o SH da cisteína e impede a reação de βCys93-SNO com moléculas X-SH do solvente.Reimpresso com permissão de Stamler, J. S., Jia, L., Eu, J. P., McMahon, T. J., Demchenko, I. T., Bonaventura, J., Gernert, K. e Piantadosi, C. A. Science 276:2034, 1997. Direitos auto-rais (1996) AAAS.
FIGURA 9.39Membranas basais. (a) Diagrama de membranas basais circundando vários tecidos e tipos celulares. (b) Micrografia eletrônica mostrando ultra-estrutura de matriz extracelular com membrana basal adjacente à célula epitelial (E). A lâmina lúcida (LL) e a lâmina densa (LD) ou lâmina basal da membrana basal são mostradas. Abaixo da lâmina densa está o estroma da matriz extracelular. Barra representa 100 nm.Parte (a) reimpressa com permissão de Kalluri, Nature Rev. Cancer 3:422, 2003. Direitos autorais (2003) Nature. Parte (b) reimpressa com permissão de Bosman, F. T. e Stamenkovic, I. J. Pathol. 200:423, 2003. Direitos autorais (2003) Pathological Society of Great Britain and Ireland. Reproduzido com permissão. Permissão dada por John Wiley & Sons, Ltd. em nome de PathSoc.
FIGURA 9.40Estrutura molecular da lâmina basal. As moléculas de laminina e de colágeno tipo IV se auto-associam para formar estruturas em rede em forma de folha, que são ligadas pelas proteínas nidogem/entactina e pelo proteoglicano de heparam sulfato perlecam. (a) Síntese de proteínas de membrana basal pela célula. (b) Montagem da rede de laminina iniciada por ligação de laminina a receptores de superfície celular. (c) Montagem da rede de colágeno tipo IV e associação de proteínas ligando os esqueletos de laminina e colágeno tipo IV. (d) Rede de colágeno tipo IV unida por interações dos domínios 7S e NC1 e com entactina/nidogem (En) fazendo ponte entre redes de colágeno e laminina (Ln).Partes (a) – (c) reimpressas com permissão de Kalluri, R. Nature Rev. Cancer 3:422, 2003. Direitos autorais (2003) Nature. Parte (d) reimpressa com permissão de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Direitos autorais (1994) Elsevier.
FIGURA 9.41Estrutura de laminina (isoforma 1). (a) Estrutura diagramática de laminina-1. As cadeias α, β e γ da laminina formam cada uma um braço curto da estrutura cruciforme. O braço longo é formado por uma estrutura espiralada-enrolada de regiões de α-hélice de todas as três cadeias com a cadeia de α-laminina estendendo-se para forma para formar o domínio globular COOH-terminal. Os braços curtos são compostos de domínios globulares, que são separados por repetições tipo fator de crescimento epidérmico (tipo-EGF). (b) Réplica de sombreamento rotatório de contraste invertido com glicerol da molécula de laminina. NH2-terminal da cadeia α e domínio globular (G) da cadeia α indicados.Parte (a) redesenhada com base em figura de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Part (b) reimpressa com permissão de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Direitos autorais (1994) Elsevier.
FIGURA 9.42Formação da estrutura em rede de laminina. (a, b) Interações de ligações entre braços curtos de diferentes moléculas de laminina levam a estruturas em rede em forma de folhas. Entactina é mostrada ligada a um braço curto de laminina, mas não é necessária para polimerização de laminina em rede. Acredita-se que o braço longo fique livre para participar de outras interações. (c) Rede de laminina vista por réplicas em platina com ângulo alto.Figuras de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Partes (a) e (b) redesenhadas e Parte (c) reimpressa com permissão. Direitos autorais (1994) Elsevier.
FIGURA 9.43Diagrama do protômero de colágeno tipo IV composto por três cadeias polipeptídicas. A linha superior mostra a localização, na estrutura primária, da região NC1, a região da tripla-hélice central (TH) com a estrutura su-per-helicoidal, a região 7S N-terminal e o peptídeo sinal (S). Barras pretas, linhas e caixas mostram localizações das múltiplas interrup-ções da seqüência Gly-X-Y nas cadeias de colágeno tipo IV.Redesenhado de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Direitos autorais (1994) Elsevier.
FIGURA 9.44Formação da estrutura em rede de colágeno tipo IV. (A) Representação esquemática da formação da rede. (a) Molécula de colágeno tipo IV protômero é formada a partir de três cadeias polipeptídicas (vermelho, púrpu-ra e azul) no retículo endoplasmático e complexo de Golgi da célula. Cadeias de colágeno são alinhadas com suas regiões 7S NH2-terminais em uma extremidade e regiões NC1 COOH-terminais na outra extremidade. (b) Dois protômeros são unidos por seus domínios NC1 para formar um dímero de protômeros (cada protômero tem três cadeias, de modo que interação é entre domínios NC1 de seis cadeias. (c) Dois dímeros de protômeros ligam-se por regiões 7S. (d) Polimerização de tetrâmeros de protômeros forma rede de colágeno tipo IV. (B) Redes de colágeno tipo IV vistas por uma réplica em ângulo alto de membrana basal amniótica observada in situ,Parte (A) redesenhada de Kalluri, R. Nature Rev. Cancer 3:422, 2003. Parte (B) reimpressa com permissão de Yurchenco, P. D. e Ruben, J. Cell. Biol. 105:2559, 1987. Modifi cado de Yurchenco, P. D. Assembly of basement membrane networks. Em: P. D. Yurchenco, D. E. Birk e R. P. Mecham (Eds.), Extracellular Matrix Assembly and Structure. New York: Academic Press, 1994, p. 351. Direitos autorais (1994) Elsevier.
FIGURA 9.45Estrutura de nidogem. Estrutura esquemática da molécula de entactina/nido-gem-1. Os sítios potenciais para ligação de cálcio são marcados por asteriscos. Sítios de ligação de colágeno tipo IV e proteoglicano ficam no domínio G2, e o sítio de ligação de laminina fica no domínio G3.Redesenhado de Erickson, A. C. e Couchman, J. R. J. Histochem. Cytochem. 48:1291, 2000.
FIGURA 9.46Detalhes moleculares das interações moleculares entre nidogem e laminina. (a) Diagrama de fita do complexo hélice-β do nidogem do domínio G3 com módulos LE3-5 de laminina. Fitas β são numeradas na folha-β da hélice. (b) Vista da interação de nidogem com domínio LE4 de laminina e a porção adjacente do domínio LE3 da laminina. Porções do esqueleto do nidogem como Cα-traço e cadeias laterais formando a interação são mostrados em dourado. Linhas traceja-das verdes mostram pontes de hidrogênio.Reimpresso com permissão de Takagi, I., Yang, Y., Lu, J., Wang, H. e Springer, T. A. Nature 424:969, 2003. Direitos autorais (2003) Nature.
FIGURA 9.47Interações de nidogem e o proteoglicano perlecam. Setas mostram interações de ligações domínio-domínio entre as proteínas principais na supra-estrutura da membrana basal.Redesenhado de Kalluri, R. Nature Rev. Cancer 3:422, 2003. FIGURA 9.49Organização modular de domínios do esqueleto protéico do perlecam. Organização em domínios do perlecam de camundongo. A ligação de cadeias de heparam sulfato (HS) é indicada. Os domínios presentes são SEA (homólogo ao domínio encontrado em proteína de esperma de ouriço-do-mar, enteroquinase e agrim); LA (homólogo ao domínio do receptor de LDL tipo A); L4 (homólogo ao domínio IV de laminina); IG (domínio tipo imunoglobulina); LE (homólogo ao domínio tipo fator de crescimento epidérmico de laminina); EG (tipo fator de crescimento epidérmico); e LG (homólogo ao domínio tipo-G de laminina).Redesenhado de Hopf, M., Göhring, W., Kohfeldt, E., Yamada, Y. e Timpl, R. Eur. J. Biochem. 259:917-925 (1999) e Kvansakul, M., Hopf, M., Ries, A., Timpl, R. e Hohenester, E. EMBO J. 20:5342, 2001.
FIGURA 9.50Complexos protéicos interconectados formam redes celulares. Diagrama mostra rede de complexos protéicos conectados encontrados em uma análise parcial do proteoma de E.coli e de levedura (proteínas expressas). O número de diferentes proteínas em um complexo varia de 2 a 83 (número médio é 12) por complexo. Linhas ligam complexos nos quais a mesma proteína foi encontrada em ambos os complexos. Os complexos mais conectados são localizados mais no centro, e complexos com pelo menos 50% de suas proteínas ortólogas a proteínas humanas são mostrados em tamanho dobrado.Complexos são codificados em cores para suas funções: cinza bem escuro, ciclo celular; cinza, sinalização; cinza- escuro, transcrição, manutenção do DNA, estrutura de cromatina; cinza-médio, transporte de proteínas e de RNA; cinza-claro, metabolismo de RNA; cinza bem claro, síntese protéica e reciclagem; cinza mais claro, biogênese de membrana e tráfego. Painel inferior mostra exemplo de ligação de um complexo com dois outros por proteínas compartilhadas. Linhas pretas mostram interações previamente listadas no Banco de Dados de Proteínas de Levedura (YPD).Reproduzido de Gavin, A.-C., Bösche, M., Krause, R., Grandi, P., et al. Nature 415:141, 2002. Direitos autorais (2002) Nature. Figura generosamente fornecida por Dr. A.-C. Gavin