Download
1 / 76
780 likes | 1.03k Views
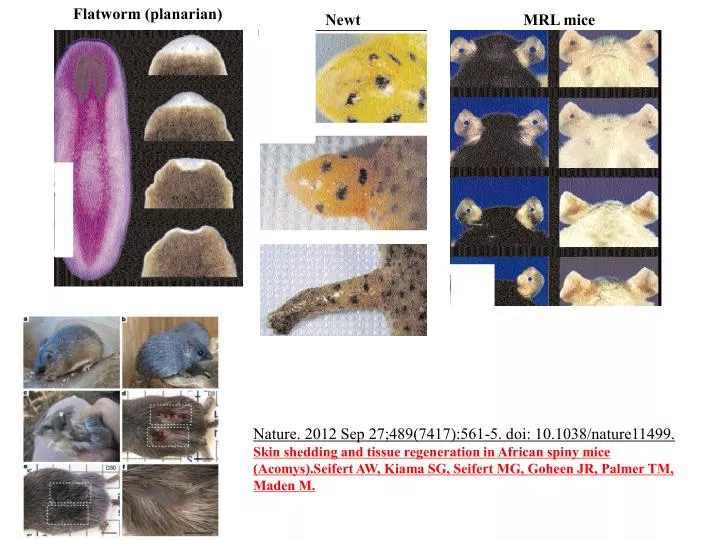
Flatworm (planarian). Newt. MRL mice. Nature. 2012 Sep 27;489(7417):561-5. doi: 10.1038/nature11499. Skin shedding and tissue regeneration in African spiny mice (Acomys).Seifert AW, Kiama SG, Seifert MG, Goheen JR, Palmer TM, Maden M. Stem Cell Development. Differentiation. functional

E N D
Flatworm (planarian) Newt MRL mice Nature. 2012 Sep 27;489(7417):561-5. doi: 10.1038/nature11499. Skin shedding and tissue regeneration in African spiny mice (Acomys).Seifert AW, Kiama SG, Seifert MG, Goheen JR, Palmer TM, Maden M.
Stem Cell Development Differentiation functional maturation Adult stem/ progenitors chronological aging Senescence PCD (apoptosis) Death ESCs
Stem Cells & Cancer • Three tumor biology puzzles: • Most tumors are of a clonal origin but tumor cells are heterogeneous. • It is very difficult to establish stable tumor cell lines from tumors. • Large numbers of established tumor cells have to be injected to re-initiate an orthotopic tumor in mice. • Key reviews: • Reya T et al. Stem cells, cancer, and cancer stem cells. Nature 414, 105-111, 2001. • Dick JE. Stem cell concepts renew cancer research. Blood 112: 4793-4807, 2008. • Visvader JE, and Lindeman GJ. Cancer Stem Cells: Current Status and Evolving Complexicities. Cell Stem Cell 10: 717-728, 2012. • 4. Tang DG. Understanding cancer stem cell heterogeneity and plasticity. Cell Res, 22(3):457-472, 2012. • 5. Magee JA, Piskounova E, & Morrison SJ. Cancer Stem Cells: Impact, Heterogeneity, • and Uncertainty. Cancer Cell 21: 283-296, 2012. • (Dean Tang, Basic Concepts of Tumor Biology, Oct 31, 2012)
Stem Cells & Cancer 1. Characteristics & Definition 2. SC Identification 3. SC Niche & Plasticity 4. SCs & Cancer 5. Cancer Stem Cells (CSCs)
Stem Cells • Rare • Generally small • - Normally localized in a ‘protected’ environment called • NICHE, where they only occasionally divide. • - But they possess HIGH PROLIFERATIVE POTENTIAL • and can give rise to large clones of progeny in vitro or in • vivo following injury or appropriate stimulation. • - Possess the ability to SELF-RENEW (i.e., asymmetric • or symmetric cell division) • - Can generate (i.e., DIFFERENTIATE into) one or • multiple or all cell types (uni-, oligo-, multi-, pluri-, or • toti-potent).
symmetric SC renewal asymmetric cell division (ACD) symmetric SC commitment (differentiation) SC Committed progenitor cells Tang, Cell Res. 2012
Cell lineage development: Self-renewal, proliferation, & differentiation Differentiation Transformation probability Self-renewal LT-SC ST-SC Late progenitors Differen- tiating cells Differen- tiated cells Early progenitors Proliferation Commitment Niche Expansion Differentiation Tang, Cell Res. 2012
Embryonic Stem Cells (ESCs) • Mouse ESCs were generated early 1980s by Evans and • Martin. • mES cells are cultured on mouse fibroblast feeders • (irradiated or mitomycin C-treated) together with LIF. • .mES cells are widely used in gene targeting. • Human ES (hES) cells were first created by Jim Thomson • (Uni. Wisconsin) in 1998. • hES cells were initially cultured also on mouse fibroblast • feeders but without LIF. Now they can be maintained in • defined medium with high bFGF (100 ng/ml), activin, • and some other factors.
How can hES cells be derived? • Leftover or dead-end IVF embryos (PGD)
Primitive ectoderm Trophectoderm Primitive Endoderm A. Nagy
ES cells A. Nagy
TS cells A. Nagy
heart pancreas testis liver brain kidney A. Nagy
Other ‘embryonic’ SCs Germline Stem Cells (GSC) Cord Blood Stem Cells (CB-SC) • Derived from umbilical cord • Primarily blood stem cells • Also contain mesenchymal stem cells that can differentiate • into bone, cartilage, heart muscle, brain, liver tissue etc. • *CB-SC could be stimulated to differentiate into neuron, • endothelial cell, and insulin-producing cells
Stem Cells & Cancer 1. Characteristics & Definition 2. SC Identification 3. SC Niche & Plasticity 4. SCs & Cancer 5. Cancer Stem Cells (CSCs)
Functional Assays of Stem Cells (Candidate) Stem Cells Stem Cells in situ (Xeno)Transplantation Lineage tracing
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assay Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
Hematopoietic stem/progenitor cell lineages Lin-CD34+CD38-CD45RA-Thy1+RholoCD49f+ (Notta F…..Dick JE. Science 333, 218-221, 2011) Lin-Sca1+ckit+CD150+CD48- (20%-50% such mouse BM cells are SCs) (~1:5,000 or 0.02%; lifetime self-renewal) (~1:1,000 or 0.1%; self-renewal for 8 wks) (No self-renewal) Passegué, Emmanuelle et al. (2003) Proc. Natl. Acad. Sci. USA 100, 11842-11849 Till JE & McCulloch EA. A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat. Res 14, 213-222, 1961.
(Nestin) (PDGFRa) (Mash-1) (NeuM) (Pax6) (A2B5) (GFAP) (MBP) (NG2)
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assay Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
Till JE & McCulloch EA. A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat. Res 14, 213-222, 1961. [The earliest report in which putative stem cells were identified by their ability to retain labeled radionucleotides for long period of time] Cotsarelis G, Cheng SZ, Dong G, Sun TT & Lavker RM. Existence of slow-cycling libmal epithelial basal cells that can be preferentially stimulated to proliferate: Implications on epithelial stem cells. Cell 57, 201-209, 1989. Cotsarelis G, Sun TT, & Lavker RM. Label-retaining cell reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell 61, 1329-1337, 1990.
LRCs in the Bulge & BM ARE Stem Cells Tumbar T et al., Defining the epithelial stem cell niche in skin. Science 303, 359-363, 2004. Blanpain, C., et al., Self-renewal, multipotency, and the existence of two cell populations in an epithelial stem cell niche. Cell 118, 635-648, 2004. Fuchs et al., Cell 116, 769, 2004 Fuchs E: The tortoise and the hair: Slow-cycling cells in the stem cell race. Cell 137, 811-819, 2009. Fuchs E & Horsley V. Ferreting out stem cells from their niches. Nat Cell Biology 13: 513-518, 2011. Wilson A et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell 135, 1118-1129, 2008. Foudi A et al. Analysis of histone 2B-GFP retention reveals slowly cycling hematopoietic stem cells. Nat. Biotechnol. 27, 84-90, 2009.
Not All Stem Cells Are Slow-Cycling ‘…. and even for the ones that do, approximately only 5-6 divisions of the label-retaining stem cell or its progeny can be monitored after a pulse-chase before the label has diluted out to the point where it can no longer be traced’. Fuchs E & Horsley V. Ferreting out stem cells from their niches. Nat Cell Biology 13: 513-518, 2011. Li L & Clevers H. Co-existence of quiescent and active adult stem cells in mammals. 327, 542-545, 2010. Not All Slow-Cycling Cells Are Stem Cells
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assays Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
E Rheinwald JG & Green H. Serial cultivation of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 6, 331-343, 1975. Sun TT Cell 9, 511-521, 1976 Nature 269, 489-493, 1977 Cell 14, 469-476, 1978 Fuchs E Cell 19, 1033-1042, 1980 Cell 25, 617-625, 1981 Barrandon Y PNAS 82, 5390-4, 1985 Cell 50, 1131-1137, 1987 Rice RH Cell 11, 417-422, 1977 Cell 18, 681-694, 1979 Watt FM JCB 90, 738-742, 1981
Anchorage- independ. survival Plating efficiency Clonogenic ‘In-gel’ assays (plate cells at low density) Prolif. potential Prolif. Clonal *Plate cells at clonal density (50-100 cells/well in 6-well plate or 10-cm dish or T25 flask) ‘On-gel’ assays (plate at low density) *Plate single cells into 96-well plates (or using flow sorting) - limiting dilution Gels: Agar Agarose Methylcellulose Matrigel Poly-HEMA fibroblasts Spheres (sphere-formation assays) Colonies (colony-formation assays) Holoclone Mero- or paraclone A colony/sphere: a 3-D structure Efficiency (%) Colony/sphere size (cell number) Colony/sphere development (tracking) Immunostaining/tumor exp. CLONAL vs CLONOGENIC ASSAYS A clone: a two-dimensional structure Cloning efficiency (CE; %) Clonal size (cell number/clone) Clonal development (tracking) Clone types
Mixing Experiments to Demonstrate the Clonality of Clones/Spheres DU145:DU145 GFP (1:1) Clonal Assay phase DU145:DU145 GFP (1:1) MC GFP DU145 RFP:DU145 GFP (1:1) MC Pastrana E, Silva-Vargas V, and Doetsch F. Eyes wide open: A critical review of sphere-formation as an assay of stem cells. Cell Stem Cell 8, 486-498, 2011
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assays Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
Goodell MA et al. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. J. Exp. Med. 183, 1797-1806, 1996. Golebiewska A, Brons NH, Bjerkvig R, and Niclou SP. Critical appraisal of the side population assay in stem cell and cancer stem cell research. Cell Stem Cell 8, 136-147, 2011.
Zhou et al., Nature Med 7, 1028, 2001 Bcrp (ABCG2) is a major mediator of the SP phenotype
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assays Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
Kastan MB et al. Direct demonstration of elevated aldehyde dehydrogenase in human hematopoietic progenitor cells. Blood 75, 1947-1960, 1990. Jones RJ et al., Assessment of aldehyde dehydrogenase in viable cells. Blood 85, 2742-46, 1995. Storms RW et al. Isolation of primitive human hematopoietic progenitors on the basis of aldehyde dehydrogenase activity. PNAS 96, 9118-9123, 1999. Alison MR, Guppy NJ, Lim SML, & Nicholson LJ. Finding cancer stem cells: Are aldehyde dehydrogenases fit for purpose? J Pathol. 222, 335-344, 2010. Ma I & Allan AL. The role of human aldehyde dehydrogenase in normal and cancer stem cells. Stem Cell Rev and Report 7, 292-306, 2011. *ALDH1A1 and ALDH3A1: Seem to be the major isozymes mediating the Aldefluor phenotype and are preferentially expressed in SC. *ALDH superfamily: 19 putatively functional genes in 11 families and 4 subfamilies. ALDH superfamily of NAD(P)+-dependent enzyme catalyzes oxidations of aldehydes to carboxylic acids.
How to identify and characterize (adult) stem cells? Marker analysis Label-retaining cells (LRC): Pulse-chase exper. Clonal/clonogenic assays Functional analysis: Side population (SP) assays Functional analysis: Aldefluor assay Cell size-based enrichment Genetic marking & lineage tracing
Fuchs E & Horsley V. Ferreting out stem cells from their niches. Nature Cell Biology 13: 513-518, 2011.
Fuchs E & Horsley V. Ferreting out stem cells from their niches. Nature Cell Biology 13: 513-518, 2011. Liver J et al., Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature 450, 56-62, 2007. Snippert HJ et al., Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lrg stem cells. Cell 143, 134-144, 2010.
Stem Cells & Cancer 1. Characteristics & Definition 2. SC Identification 3. SC Niche & Plasticity 4. SCs & Cancer 5. Cancer Stem Cells (CSCs)
Stem Cell Niche in Hair Follicles: The Bulge Moore KA & Lemischka IR. Science 311, 1880-1885, 2006
Bulge Stem Cells Tumbar et al., Science 303, 359-363, 2004; Fuchs et al., Cell 116, 769, 2004
Stem Cell Niche in Small Intestine: The Crypt Moore KA & Lemischka IR. Science 311, 1880-1885, 2006 Barker N et al., Cell Stem Cell 11: 452-460, 2012.
Stem Cell Niches in BM Moore KA & Lemischka IR. Science 311, 1880-1885, 2006 Naveiras O et al., Bone-marrow adipocytes as negative regulators of the hematopoietic microenvironment. Nature 460, 259, 2009. Mendez-Ferrer, S et al., Mesenchymal and hematopoietic stem cells form a unique bone marrow niche. Nature 466, 829-834, 2010.
Stem cell lineage Differentiated cells Progenitors/ Precursor cells Other differ. cell(s) Senescence Death (PCD) Stem cells
“Transdifferentiation” of Stem Cells *First report: Long-term cultured adult brain (stem) cells can reconstitute the whole blood in lethally irradiated mice (Bjornson et al., Science 283, 534-537, 1999). *Cells from skeletal muscle have hematopoietic potential (Jackson et al., PNAS 96, 14482-14486, 1999) and can also “differentiate” into many other cell types (Qu-Petersen, Z, et al., JCB 157, 851- 864, 2002). *CNS “SCs” can “differentiate” into muscle cells (Clarke et al., Science 288, 1660-1663, 2000; Galli et al., Nat. Neurosci 3, 986-991, 2000; Tsai and McKay, J. Neurosci 20, 3725-3735, 2000). *Vice versa, “SCs” from blood or bone marrow can “transdifferentiate” into muscle (Ferrari et al., Science 279, 1528-1530, 1998; Gussoni et al., Nature 401, 390-394, 1999), hepatocytes (Petersen et al., Science 284, 1168-1170, 1999; Lagasse et al., Nat Med 6, 1229-1234, 2000), cardiac myocytes (Orlic et al., Nature 410, 701-705, 2001), or neural cells (Mezey et al., Science 290, 1779-1782, 2000; Brazelton et al., Science 290, 1775-1779, 2000). *Bone marrow appears to contain two distinct SCs: the HSC and MSC. A single HSC could contribute to epithelia of multiple organs of endodermal and ectodermal origin (Krause et al., Cell 105 369-377, 2001). MSC, on the other hand, can adopt a wider range of fates (endothelial, liver, neural cells, and perhaps all cell types) (Pittenger et al., Science 284, 143-146, 1999; Schwartz et al., JCI 109, 1291-1302, 2002; Jiang et al., Nature 418, 41-49, 2002). *Pluripotent “SCs” have also been isolated from skin that can “differentiate” into neural cells, epithelial cells, and blood cells (Toma et al., Nat Cell Biol. 3, 778-784, 2001) *Highly purified adult rat hepatic oval “stem’ cells, which are capable of differentiating into hepatocytes and bile duct epithelium, can “trans-differentiate” into pancreatic endocrine hormone- producing cells when cultured in a high glucose environment (Yang et al., PNAS 99, 8078- 8083, 2002)
De-differentiation: Cell-cycle re-entry *Many ‘post-mitotic’ cells such as hepatocytes, endothelial cells, and Schwann cells have long been known to retain proliferative (progenitor) potential. *Dedifferentiation is a genetically regulated process that may ensure a return path to the undifferentiated state when necessary (Katoh et al., PNAS 101, 7005, 2004). *Regeneration of male GSC by spermatogonial dedifferentiation in vivo (Brawley and Matunis, Science 304, 1331, 2004). *Conversion of mature B cells into T cells by dedifferentiation to uncommitted progenitors (Nature 449, 473-477, 2007). *During Salamander limb regeneration, complete de-differentiation to a pluripotent state is not required – Progenitor cells in the blastema keep a memory of their tissue origin (Nature 460, 60-65, 2009). *Epigenetic reversion of post-implantation epiblast to pluripotent embryonic cells (Nature, 461, 1292-1295, 2009). *Evidence for cardiomyocyte renewal in humans (Bergmann O et al., Science 324, 98-102, 2009). (Cardiomyocytes turn over at an estimated rate of ~1% per year at age 20, declining to 0.4% per year at age 75. At age 50, 55% of human cardiomyocytes remain from birth while 45% were generated afterward. Over the first decade of life, cardiomyocytes often undergo a final round of DNA synthesis and nuclear division without cell division, resulting in ~25% of human cardiomyocytes being binucleated.) *Neuregulin 1/ErbB4 signaling induces cardiomyocyte proliferation and repair of heart injury (Bersell et al., Cell, 138, 257-270, 2009). *MafB/c-Maf deficiency enables self-renewal of differentiated functional macrophages (Aziz A, et al., Science 326, 867-871, 2009).
Pancreatic b-cells: Interesting insulin-producing cells *Insulin-producing b-cells in adult mouse pancreas can self-duplicate during normal homeostasis as well as during injury (Dor et al., Nature 429, 41, 2004). *In vivo reprogramming of adult pancreatic exocrine cells to b cells using 3 TFs (Ngn3, Pdx1, and Mafa), suggesting a paradign for directing cell reprogramming without reversion to a pluripotent cell state (Zhou et al., Nature 455, 627-632, 2008). *In response to injury, a population of pancreatic progenitors can generate glucagon-expressing alpha cells that then transdifferentiate (with ectopic expression of Pax4) into beta cells (Collmbat et al, Cell 138, 449-462, 2009). *Conversion of adult pancreatic a-cells to b-cells after extreme b-cell loss (Nature 464, 1149-1154, 2010).