Download

1 / 66
700 likes | 1.41k Views
Nucleotide Metabolism. UNIT IV: Nitrogen Metabolism. Overview. Nucleotides are essential for all cells DNA and RNA synthesis/protein synthesis/cell proliferation Carriers of activated intermediates in synthesis of some CHO’s, lipids and proteins

E N D
Nucleotide Metabolism UNIT IV: Nitrogen Metabolism
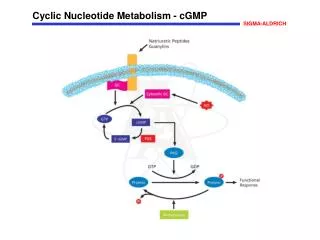
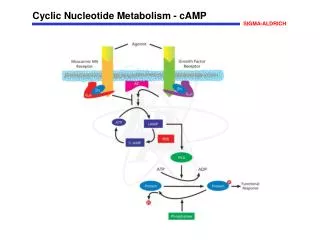
Overview • Nucleotides are essential for all cells • DNA and RNA synthesis/protein synthesis/cell proliferation • Carriers of activated intermediates in synthesis of some CHO’s, lipids and proteins • Structural components of several essential coenzymes, e.g., CoA, FAD, NAD+, NADP+ • cAMP and cGMP serve as second messengers in signal transduction • Energy currency • Regulatory compounds for many pathways of intermediary metabolism • Purine and pyrimidine bases can be synthesized de novo, or obtained through salvage pathways
Nucleotide structure • Nucleotides are composed of a nitrogenous base (purine/pyrimidine), a pentose, and 1, 2 or 3 phosphate groups. Purine and pyrimdine structures • Both DNA and RNA contain purine bases: adenine (A) and guanine (G). • Both DNA and RNA contain cytosine (C) • DNA contains thymine (T), whereas RNA contains uracil (U). T and U differ by one methyl group
Unusual bases are occasionally found in some species of DNA and RNA e.g., in some viral DNA, tRNA Base modifications include methylation, hydroxymethylation, glycosylation, acetylation, or reduction May aid in recognition by specific enzymes/proteins, or protect from degradation by nucleases
Nucleosides • Pentose sugar + N-base = nucleoside (ribonucleoside/ deoxyribonucleosides). • Ribonucleosides of A, G, C and U = adenosine, guanosine, cytidine, uridine • Deoxyribonucleosides have the added prefix “deoxy” • Carbon and nitrogen atoms in base and sugar are numbered separately
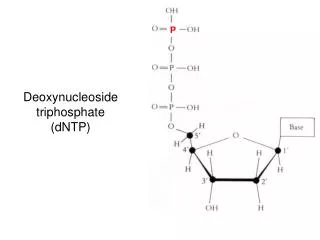
Nucleotides • Nucleotides are mono-, di-, or triphosphate esters of nucleosides. The 1st phosphate group is attached by an ester linkage to the 5`-OH of pentose. This compound = nucleoside 5`-phosphate or 5`-nucleotide • If one phosphate is attached, the structure = nucleoside monophosphate (NMP) e.g., AMP, CMP • If a 2nd or 3rd phosphate is added, a nucleoside diphosphate (e.g., ADP) or triphosphate (e.g., ATP) • The 2nd and 3rd phosphates are connected by a ”high-energy” bond. • Phosphate groups give negative chrges to nucleotides, and cause DNA and RNA to be referred to as “nucleic acids”.
Synthesis of Purine Nucleotides • Atoms of purine ring are contributed by: Asp, Gly and Gln/ CO2/ and N10-formyl-tetrahydrofolate. Ring is constructed by reactions that add donated carbons and nitrogens to preformed ribose-5-P. Sources of individual atoms in the purine ring
A. Synthesis of 5-phosphoribosyl-1-pyrophosphate (PRPP) • PRPP is an “activated pentose”, participates in synthesis of purines and pyrimidines, and in salvage of purine bases • Synthesis of PRPP from ATP and ribose-5-P is catalyzed by PRPP synthetase (ribose phosphate pyrophosphokinase). • The enz. is activated by (Pi) and inhibited by purine nucleotides (end-product) • Sugar in PRPP is ribose, so end products of de novo purine synthesis ribonucleotides, when deoxyribonucleotides needed for DNA syhtnesis, ribose is reduced
B. Synthesis of 5`-phosphoribosylamine • Amide group of Gln replaces the pyrophosphate group attached to C1 of PRPP • The enz, glutamine:phosphoribosyl pyrophosphate amidotransferase, inhibited by purine 5` nucleotides AMP, GMP, IMP (end products). • This is committed step in purine biosynthesis • The rate of rxn is also controlled by Gln and PRPP conc. (intracellular PRPP conc is normally << Km for the amidotransferase, i.e., small changes in [PRPP] cause proportional change)
C. Synthesis of inosine monophosphate, the “parent” purine nucleotide • The next 9 steps in purine nucleotide biosynthesis lead to IMP (whose base is hypoxanthine). • The pathway requires 4 ATPs. Two steps require N10-formyltetrahydrofolate.
D. Synthetic inhibitors of purine synthesis • Sulfonamides inhibit growth of rapidly dividing m/o’s without interfering with human cell functions. • Methotrexate (structural analog of folic acid) used to control spread of cancer by interfering with synthesis of nucleotides (thus DNA, RNA). • Inhibitors of human purine synthesis are extremely toxic to tissues, esp. to developing structures such as in a fetus , or to cell types that replicate rapidly, including those of BM, skin, GI tract, immune system, or hair follicles. • Individuals taking such anti-cancer drugs experience adverse effects e.g., anemia, scaly skin, GI tract disturbance, immunodeficiencies, and baldness • Trimethoprim (another folate analog) has antibacterial activity as it selectively inhibits bacterial dihydrofolate reductase.
E. Conversion of IMP to AMP and GMP • This requires a 2-step energy requiring pathway • Synthesis of AMP requires GTP and that of GMP requires ATP • The 1st rxn in each pathway is inhibited by the end product. This diverts IMP to the synthesis of the purine species present in lesser amounts. • If both AMP and GMP present in adequate amounts, de novo pathway of purine synthesis is turned off at amidotransferase step. • Mycophenolic acid (MPA) is a potent, reversible, uncompetitive inhibitor of inosine monophosphate dehydrogenase in use for preventing graft rejection. Blocks de novo formation of GMP, thus depriving rapidly proliferating cells, icluding T and B cells of a key component of nucleic acids
F. Conversion of nucleoside monophosphates (NMPs) to nucleoside diphosphates (NDPs) and triphosphates (NTPs) • NDPs are synthesized from NMPs by base-specific nucleoside monophosphate kinases. These kinases do not discriminate b/w ribose or deoxyribose in the substrate. • ATP is generally the source of transferred phosphate as it is the abundant NTP • Adenylate kinase is particularly active in liver and muscle, where turnover of energy from ATP is high. Its function is to maintainan equilibrium among AMP, ADP and ATP • NDPs and NTPs are interconverted by nucleoside diphosphate kinase, an enz. with broad specificity.
G. Salvage pathway of purines • Purines from normal turnover of cellular nucleic acids, or obtained from diet and not degraded, can be reconverted into NTPs and used by the body. This is “salvage pathway” of purines. 1. Conversion of purine bases to nucleotides: - Two enz’s are involved: adenine phosphoribosyltransferase (APRT) and hypoxanthine-guanine phosphoribosyltransferase (HPRT). - Both enz’s use PRPP as source of ribose 5-p. Release of ppi makes these rxns irreversible.
2. Lesch-Nyhan syndrome: • X-linked, recessive disorder, associated with virtually complete deficiency of HPRT. • Inability to salvage hypoxanthine or guanine, from which excessive amounts of uric acid are produced. • In addition, lack of salvage pathway causes increased PRPP levels and decreased IMP and GMP levels. • So, glutamine:phosphoribosylpyrophosphate amidotransferase has excess substrate and decreased inhibitors available, and de novo purine synthesis increased. • Decreased purine reutilization and increased purine synthesis results in production of large amounts of uric acid, making Lesch-Nyhan syndrome a severe, heritable form of gout. • Patients with Lesch-Nyhan syndrome tend to produce urate kidney stones. • In addition, characteristic neurologic features of the disorder include self-mutilation and involuntary movements.
IV. Synthesis of Deoxyribonucleotides • Ribonucleotide reductase (ribonucleoside diphosphate reductase) - Is a multisubunit enzyme (2B1 + 2B2) specific for reduction of nucleoside diphosphates (ADP, GDP, CDP, UDP) to their deoxy forms. The immediate donors of hydrogen atoms needed for reduction of 2`-OH are two –SH groups on enz itself, which during rxn form a disulfide bond.
1. Regeneration of reduced enzyme. • Disulfide bond must be reduced. Source of reducing equivalents is thioredoxin- a peptide coenzyme of ribonucleotide reductase. • Thioredoxin contains two cys residues separated by two aa’s in the peptide chain. The two –SH groups of thioredoxin donate their H atoms to enz., in the process forming S-S bond. 2. Regeneration of reduced thioredoxin. • The necessary reducing equivalents are provided by NADPH,H+
B. Regulation of deoxyribonucleotide synthesis • Ribonucleotide reductase is responsible for maintaining a balanced supply of the deoxyribonucleotides required for DNA synthesis. • Regulation of the enz is complex. In addition to the single catalytic (active) site, there are sites on the enz involved in regulating its activity. • Activity sites: binding of dATP to allosteric sites (known as the activity sites) on the enz inhibits the overall catalytic activity of the enz prevents reduction of any of the four NDPs. This effectively prevents DNA synthesis, and explains the toxicity of increased levels of dATP seen in conditions such as adenosine deaminase deficiency.
Substrate specificity sites: binding of dNTPs to additional allosteric sites (known as the substrate specificity sites) on the enz regulates substrate specificity, causing an increase in the conversion of different species of ribonucleotides to deoxyribonucleotides as they are required for DNA synthesis. E.g., dTTP binding at the specificity sites causes a conformational change that allows reduction of GDP to dGDP at the catalytic site.
Hydroxyurea • The drug, hydroxyurea destroys the free radical required for enzymic activity of ribonucleotide reductase, and thus inhibits the generation of substrates for DNA synthesis. • Hydroxyurea has been used in the treatment of cancers such as chronic myelogenous leukemia. Hydroxyurea is also used in the treatment of sickle cell disease however, the increase in fetal hemoglobin seen with hydroxyurea has not been linked to its effect on ribonucleotide reductase.
V. Degradation of Purine Nucleotides • Degradation of dietary NAs occurs in the small intestine, where a family of pancreatic enzymes hydrolyzes the nucleotides to nucleosides and free bases. • Inside cells, purine nucleotides are sequentially degraded by specific enzymes, with uric acid as the end product of this pathway. [Note: Mammals other than primates oxidize uric acid further to allantoin, which, in some organisms other than mammals, may be further degraded to urea or ammonia.]
A. Degradation of dietary nucleic acids in the small intestine • Ribonucleases and deoxyribonucleases, secreted by the pancreas, hydrolyze RNA and DNA primarily to oligonucleotides. • Oligonucleotides are further hydrolyzed by pancreatic phosphodiesterases, producing a mixture of 3′- and 5′-mononucleotides. • A family of nucleotidases removes the phosphate groups hydrolytically, releasing nucleosides that may be absorbed by the intestinal mucosal cells, or be further degraded to free bases before uptake. • Dietary purines and pyrimidines are not used to a large extent for the synthesis of tissue nucleic acids. Instead, the dietary purines are generally converted to uric acid by intestinal mucosal cells. • Most of the uric acid enters the blood, and is eventually excreted in the urine.
B. Formation of uric acid [1] An amino group is removed from AMP to produce IMP, or from adenosine to produce inosine (hypoxanthine-ribose) by AMP deaminase or adenosine deaminase. [2] IMP and GMP are converted into their nucleoside forms—inosine and guanosine—by the action of 5′-nucleotidase. [3] Purine nucleoside phosphorylase converts inosine and guanosine into their respective purine bases, hypoxanthine and guanine. [Note: A mutase interconverts ribose 1- and ribose 5-phosphate]. [4] Guanine is deaminated to form xanthine. [5] Hypoxanthine is oxidized by xanthine oxidase to xanthine, which is further oxidized by xanthine oxidase to uric acid, the final product of human purine degradation. Uric acid is excreted in the urine.
C. Diseases associated with purine degradation • Gout: Gout is a disorder characterized by high levels of uric acid—the end product of purine catabolism—in blood (hyperuricemia), as a result of either the overproduction or underexcretion of uric acid. - The hyperuricemia leads to the deposition of monosodium urate crystals in the joints, and an inflammatory response to the crystals, causing first acute and then to chronic gouty arthritis. - Nodular masses of monosodium urate crystals (tophi) may be deposited in the soft tissues, resulting in chronic tophaceous gout. - Formation of uric acid stones in the kidney may also be seen. [Note: Hyperuricemia is typically asymptomatic and does not lead to gout, but gout is preceded by hyperuricemia.] - The definitive diagnosis of gout requires aspiration and examination of synovial fluid from an affected joint (or material from a tophus) using polarized light microscopy to confirm the presence of needle-shaped monosodium urate crystals.
Analysis of joint fluid can help to define causes of joint swelling or arthritis, such as infection, gout, and rheumatoid disease
Underexcretion of uric acid: In the vast majority of patients, the hyperuricemia leading to gout is due to underexcretion of uric acid. - Underexcretion can be primary, due to as-yet-unidentified inherent excretory defects, or secondary to known disease processes that affect how the kidney handles urate, e.g., lactic acidosis, and to environmental factors such as the use of drugs, e.g., thiazide diuretics, or exposure to lead (saturnine gout).
b. Overproduction of uric acid: A less common cause of gout is hyperuricemia from the overproduction of uric acid. - Primary hyperuricemia is, for the most part, idiopathic. However, several identified mutations in the X-linked PRPP synthetase gene result in the enzyme having an increased Vmax for the production of PRPP, a lower Km for ribose 5-phosphate, or a decreased sensitivity to purine nucleotides—its allosteric inhibitors. - In each case, increased availability of PRPP increases purine production, resulting in elevated levels of plasma uric acid. - Lesch-Nyhan syndrome also causes hyperuricemia as a result of the decreased salvage of hypoxanthine and guanine, and the subsequent increased availability of PRPP.
- Secondary hyperuricemia is typically the consequence of increased availability of purines, e.g., in patients with myeloproliferative disorders or who are undergoing chemotherapy and so have a high rate of cell turnover. - Hyperuricemia leading to gout can also be the result of seemingly unrelated metabolic diseases, such as von Gierke disease or fructose intolerance.
Secondary hyperuricemia resulting in gout can also be caused by excessive consumption of ethanol and by diet. • Individuals with a history of gout traditionally have been cautioned about consuming foods such as organ meats, anchovies, sardines, and legumes, as these foods were thought to be associated with increased risk of a gouty attack. • Recent studies on the relationship between diet and gout have confirmed that a diet rich in meat and seafood is associated with increased risk. In addition, a diet rich in low-fat dairy products was shown to be associated with a decreased risk.
c. Treatment of gout: Acute attacks of gout are treated with anti-inflammatory agents. Colchicine, steroidal drugs such as prednisone, and nonsteroidal drugs such as indomethacin are used. - [Note: Colchicine depolymerizes microtubules, thus decreasing the movement of neutrophils into the affected area. Like the other anti-inflammatory drugs, it has no effect on uric acid levels.] - Long-term therapeutic strategies for gout involve lowering the uric acid level below the saturation point, thereby preventing the deposition of urate crystals.
- Uricosuric agents, such as probenecid or sulfinpyrazone, that increase renal excretion of uric acid, are used in patients who are “underexcretors” of uric acid. - Allopurinol, an inhibitor of uric acid synthesis, is used in patients who are “overproducers” of uric acid. Allopurinol is converted in the body to oxypurinol, which inhibits xanthine oxidase, resulting in an accumulation of hypoxanthine and xanthine —compounds more soluble than uric acid and, therefore, less likely to initiate an inflammatory response. In patients with normal levels of HGPRT, the hypoxanthine can be salvaged, thus reducing the levels of PRPP and, therefore, de novo purine synthesis.
2. Adenosine deaminase (ADA) deficiency: • ADA is expressed in the cytosol of all cells, but, in humans, lymphocytes have the highest activity of this enzyme. • A deficiency of ADA results in an accumulation of adenosine, which is converted to its ribonucleotide or deoxyribonucleotide forms by cellular kinases. • As dATP levels rise, ribonucleotide reductase is inhibited, thus preventing the production of all deoxyribose-containing nucleotides.
Consequently, cells cannot make DNA and divide. In its most severe form, this autosomal recessive disorder causes a type of severe combined immunodeficiency disease, involving a decrease in both T cells and B cells. • It is estimated that in the United States, ADA deficiency accounts for approximately 14% of all cases of SCID. • Treatment requires either bone marrow replacement or enzyme replacement therapy. Without appropriate treatment, children with this disorder usually die by the age of two.