Download

1 / 27
270 likes | 285 Views
Explore the intricate processes of photosynthesis, including the light reactions on thylakoid membranes and the Calvin Cycle. Learn about Photosystem I and Photosystem II, electron transport, ATP and NADPH production, and how plants convert light energy into chemical energy for growth. Discover how oxygen is released during photosynthesis and the significance of this essential process. Delve into the dark reactions driven by ATP and NADPH to produce carbohydrates.

E N D
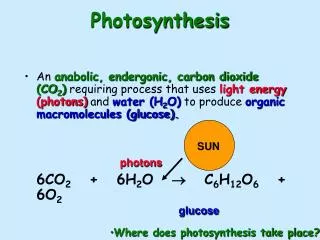
Photosynthesis Photosynthesis involves two sets of reactions: the light reactions and ‘dark’ reactions that are otherwise called the Calvin cycle. The Light Reactions These reactions occur on the thylakoid membranes of chloroplasts. There are two photosystems involved, named, logically enough, Photosystem I and Photosystem II. Each of these photosystems contains proteins complexed with cholorophyll pigments, and photosystem II also contains carotenoids. The chlorophyll and carotenoids are organized into light harvesting complexes. They trap photons.
The energy of the trapped photon excites a chlorophyll a molecule and, through what is called resonance, that energy is transferred to a Photosystem reaction centre. Either directly (if the photon excited Photoystem I) or indirectly via electron transport (if the photon excited Photosystem II), light energy is converted into electron energy that is used in electron transport to NADP to reduce it to NADPH, splitting water and releasing an electron and oxygen, or to phosphorylate ADP to ATP.
Showing both photosystems I and II…in addition to reducing NADP to NADPH (photosystem I) ATP is produced when light excites photosystem II.
The equivalent diagram from the text looks slightly different, but expresses exactly the same reaction system…
The diagram on the two previous slides showed the excitation of Photosystem II caused P680 to become oxidized (lose an electron). That electron is replaced by one from water (as part of splitting water into hydrogen and oxygen). That electron is passed through a series of carriers, losing some energy in each step. The labeled sequence of acceptors are Pg (plastiquinone), a cytochrome complex and plastocyanin. That energy is captured (partly) in the formation of an ATP molecule from ADP (called photophosphorylation). The electron is eventually passed to an oxidized P700 of photosystem I. When P700 was itself excited by light, it was oxidized, and passed an electron through a series of acceptors, with the energy used to reduce NADP to NADPH. This whole process is non-cyclic photophosphorylation.
Just to make it all more complicated, photosystem I can function independent of photosystem II, in cyclic photophosphorylation. This sequence of electron transfer produces ATP directly, rather than forming NADPH.
The capture of photon energy occurs on the thylakoid membranes that form the characterstic grana stacks within the chloroplasts. Once more, your text has an excellent figure…
The entire photochemical reaction occurs incredibly quickly. From the initial capture of the photon to the formation of NADPH and or ATP, the process is complete in a few picoseconds (one picosecond = 10-12 seconds). What is particularly important to us about photosynthesis is that it is the process that produces/releases free O2 into the atmosphere. There was no free oxygen formed before about 2-2.5 billion years ago, when photosynthesis first evolved in some bacteria. Even then, there was not enough formed to result in free oxygen in the atmosphere. That only occurred about 1.5 billion years ago with an increase in the number of photosynthetic organisms and a saturation of oxygen absorbers like iron-containing (and magnesium- and other metal ion containing) minerals.
The Dark Reactions The energy captured in ATP and NADPH is used to drive the chemical reactions of the Calvin-Benson cycle. Melvin Calvin, from the University of California, won a Nobel Prize for the ‘discovery’ and description of the dark reactions (meaning not light requiring) of photosynthesis. Here’s one diagram of the process. The molecules involved: RuBP – ribulose biphosphate PGA – phosphoglyceric acid PGAL – phosphoglyceraldehyde rubisco – ribulose biphosphate carboxylase
The steps of the Calvin cycle are: • Fixation of CO2 by enzymatically adding a carbon to ribulose 1,5 biphosphate. The enzyme is rubisco (ribulose biphosphate carboxylase. Rubisco is the most common protein in photosynthetic plants, representing from 1/8 to 1/4 of total leaf protein. • The 6-carbon molecule formed is unstable, and very rapidly splits into two 3-carbon molecules of phosphoglyceric acid (PGA). • PGA is modified enzymatically (with the energy input from one NADPH and one ATP from the light reactions) into two molecules of glyceraldehyde phosphate (PGAL). Most of the PGAL (10 out of every 12) is used to regenerate RuBP. That makes the series of reactions cyclic.
4. The other two PGAL are re-combined enzymatically to form a 6-carbon sugar, fructose 1,6 biphosphate. That sugar molecule is converted rapidly to glucose, which is, in turn, converted into sucrose or starch.
There are limitations to the amount of carbon fixed into carbohydrates by photosynthesis and the Calvin-Benson reactions: 1. The process may be limited by the amount of light available, and in the basic C3 system by the balance between photosynthesis and photorespiration. What is frequently measured for plants is the light compensation point. That is the light level at which photosynthesis and photorespiration are in balance, and there is 0 net fixation of carbon.
In C3 plants photorespiration competes with photosynthesis for ribulose biphosphate. In photosynthesis a carbon is ‘added’ and molecules of 3-phosphogycerate are formed. In photorespiration the same enzyme (rubisco) binds oxygen to ruBP. It should be obvious that high O2 concentration in a leaf leads to higher rates of photorespiration. It is not obvious, but true that higher temperature also promotes photorespiration. Photosynthesis releases free oxygen. Photorespiration, logically enough, releases CO2 as its ‘waste’ product, but without formation of ATP or NADPH. Complete photorespiration involves not only the chloroplasts, but also peroxisomes and mitochondria.
Photorespiration occurs in C3 plants, but not in C4 plants. On a hot day, photo-respiration may cost a C3 plant as much as 50% of fixed carbon, at a high energy cost. On cooler days (or in cooler climates) when photo-respiration is low or unlikely to occur, C3 plants are more efficient (expend less energy) to fix CO2.
Species have evolved physiological differences to parallel their growth strategies. The light compensation point may change seasonally (in part by changing amounts of photosynthetic pigments and rubisco), and the maximum rate of photosynthesis may also change. The text table presents results of a study of forest understory herbs that grew (and flowered) in spring (Allium tricoccum) when there was little else in full leaf to shade the plant, in summer when light competition is most severe (Viola pubescens), and a Saxifrage that grows right through to autumn (Tiarella cordifolia).
Since light competition is so severe, it affects not just interactions between species and individuals within a species, but even the evolution of growth patterns within individuals. Plants present their leaves so as to minimize the amount of self-shading occurring among those leaves. How? Among the taxonomic characteristics used to identify plants are whether their leaves are opposite or adjacent, whether leaves occur above one another or in whorls, whether the leaves are entire or various more complicated shapes with more-or-less finger-like projections. Some tropical plant leaves even have holes in them. Each of these is a portion of the adaptive scheme to maximize light capture efficiency and minimize the cost of maintaining the light capture apparatus.
Leaves in whorls Opposite leaves Alternate leaves Leaves may even have different shapes on the same plant where they occur in open sun versus in the shade. A related question is what part of sunlight does the photosynthetic apparatus absorb? Not all light is photosynthetically active radiation (or PAR). Leaves look green because that part of the spectrum is not absorbed, but is reflected, so that’s what we see.
2. There may be a limitation in the amount of photo-synthetically active radiation available. Only two forms of chlorophyll are important in photosynthesis – a (the photon capture pigment for photosynthesis) and b, and they have radically different absorption spectra… Latitude, and the thickness of atmosphere that light has to penetrate to reach the earth’s surface, also make a difference in the relative intensities of different parts of the light spectrum.
3. Under some circumstances there can also be a limit in the amount or rate at which leaves can take in CO2. In part this is due to the limits to diffusion rate of CO2 through the stomates of the leaf. One part of the limit is set by leaf conductance, which is the rate of CO2 movement at a given difference between ambient CO2 in the atmosphere around the leaf and intercellular concentration of CO2 within the leaf. Your text goes into further detail, subdividing the binding process into compartments that include what goes on inside the cells where photosynthesis occurs… Another part of the limit is set by the binding coefficient for CO2 by the rubisco enzyme, at least for plants with C3 photosynthesis.
There can even be significant variation in photosynthetic rate across the area of a single leaf. This is logical based on shading, leaf thickness, and spatial variation in water stress leading to variation in stomatal opening…
Alternatives to C3 photosynthesis The Calvin-Benson cycle is universal in photosynthetic plants. However, as you already know, there are alternatives in carbon fixation. In C4 photosynthesis, the initial carbon fixation step uses PEP carboxylase to attach a carbon from CO2 to phosphoenol- pyruvate, a 3-carbon molecule, to form a 4-carbon molecule, oxaloacetate. There are then a cycle of reactions during which a CO2 is passed to the Calvin cycle. Note the location of these steps within the leaf. This mode of carbon fixation is called the Hatch-Slack pathway.
Carbon fixation only occurs in the mesophyll, using PEP carboxylase, which has a far higher binding affinity for CO2. The 4-C products of fixation are passed to the bundle sheath, where the fixed carbon is moved into the Calvin-Benson pathway.
C3 and C4 leaf structures are different, as well. C4 plants have what is called the Krantz anatomy. Chloroplasts are more densely packed in a ring of mesophyll cells directly surrounding the bundle sheath, which is frequently protected by high lignin and silica content in the C4 anatomy. Mesophyll cells are more loosely packed and chloroplasts more uniformly distributed in C3 plants.
Many other differences follow from the difference in carbon fixation and anatomy. The absence of photorespiration in C4 plants (rubisco is mostly all in cells buried deep in the leaf surrounding the bundle sheath) means that net photosynthesis will continue to increase with leaf temperature to much higher temperatures in C4s. The concentration of CO2 in the bundle sheath of C4s is far higher than in C3s.
Because PEP carboxylase’s binding affinity for CO2 is high, C4 plants have the ability to continue to fix carbon for a while even when their stomates are closed. That makes the water use efficiency of C4s much higher than that for C3s. That same affinity means that far less PEP carboxylase protein is needed than the requirement for rubisco in C3s. That results in a far greater nitrogen use efficiency, as well. All these things, mostly resulting from the limitations to using rubisco to initially fix carbon, mean that C4 plants can continue to increase photosynthesis as light intensity increases to much higher light levels than will saturate C3 plants.
What about CAM (Crassulacean Acid Metabolism) plants? CAM uses the same pathways as C4, but with much different localization and adaptations. CAM plants are mostly found in hot environments and have thick, succulent leaves. In CAM plants rubisco is distributed throughout the leaf mesophyll, not just in the bundle sheath. Therefore, they can’t afford to have their stomates open during the day. They fix carbon at night with stomates open – it’s cooler and typically more humid. They have to ‘store’ enough carbon overnight to carry them through the day, when the light reactions occur. Since stomates are then closed, the ‘release’ of carbon fixed by PEP carboxylase overnight means that rubisco works in a high CO2 – low O2 environment, minimizing any chance of photorespiration.
So, the C4 system separates carbon fixation and rubisco spatially (mesophyll versus bundle sheath), while CAM separates PEP carboxylase and rubisco function temporally (PEP carboxylase-driven carbon fixation at night versus rubisco function with stomates closed during daylight).