Download

1 / 58
580 likes | 855 Views
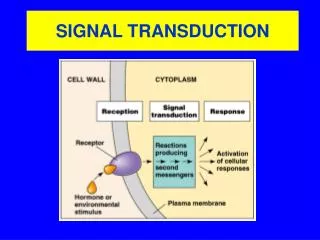
Chapter 14 Signal-Transduction Pathway. Signal-transduction circuits in biological systems have molecular on-off switches that, like those in a computer chip, transmit information when ”on”. Chemical signals are crucial to coordinating physiological responses.

E N D
Chapter 14 Signal-Transduction Pathway
Signal-transduction circuits in biological systems have molecular on-off switches that, like those in a computer chip, transmit information when ”on”.
Chemical signals are crucial to coordinating physiological responses. When organism is threatened, the adrenal glands release the hormone epinephrine, which stimulates the mobilization of energy stores and leads to improved cardiac function. After full meal, the cells in the pancreas release insulin, which stimulates the uptake of glucose from the bloodstream and leads to other physiological changes. The release of epidermal growth factor in response to a wound stimulates specific cells to grow and divide.
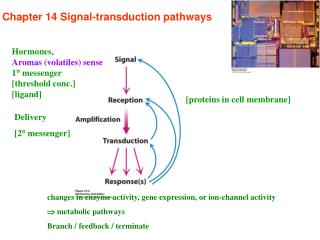
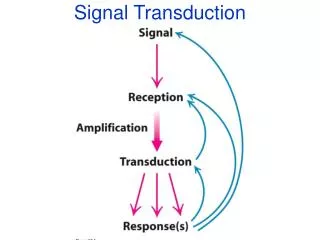
Signal transduction depends on molecular circuits • Release of the primary messenger. • - ligand 2. Reception of the primary messenger. 3. Delivery of the message inside the cell by the second messenger(cAMP, cGMP, Ca2+, IP3, DAG…). 4. Activation of effectors that directly alter the physiological response. 5. Termination of the signal.
※ The use of second messenger : • > the signal may be amplified. • > free to diffuse to other cellular compartments. • > creates both opportunities and potential problem.
14.1 Heterotrimeric G proteins transmit signals and reset themselves • Epinephrine ↔ β–adrenergic receptor(β-AR) • β–adrenergic receptor : member of 7 transmembrane helix(7TM) receptors. • 7TM receptor : initiated by signals as diverse as hormones, neurotransmitters, odorants, tastants, and protons.
- 7TM (= serpentine receptor) The single polypeptide chain “snake” through the membrane 7 times. - Rhodopsin : essential role in vision. Member of 7TM family. - 7TM family members are similar in structure to rhodopsin.
- The binding of epinephrine to β–adrenergic receptor triggers conformational changes in the cytoplasmic loops and the C-terminal region.
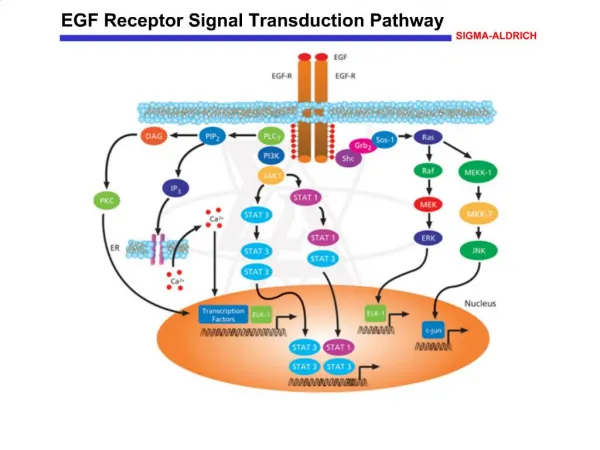
Ligand binding to 7TM receptors leads to the activation of heterotrimeric G proteins Epinephrine binding β–AR conformational change Activate G protein Stimulate adenylate cyclase Increase cAMP concentration
Hormone binding → receptor stimulate nucleotide exchange(GDP → GTP) • ※Inactivated state of G protein • GDP bound at α subunit. • Exist as heterotrimeric(αβγ). • αsubunit is p-loop NTPase(nucleotide binding) family member. • α and γ subunits are anchored to the membrane by lipid modification. • ※Activated state of G protein • GTP bound at α subunit • αsubunit dissociates from β and γ subunits.
Activated G proteins transmit signals by binding to other proteins -When G protein activated, αsubunit dissociated from βγsubunits. And αsubunit find new binding partner(Adenylate cyclase). - Adenylate cyclase : convert ATP into cAMP. 12 membrane spanning helices. 2 large cytoplasmic domains. - Result : binding of epinephrine to the receptor on the cell surface increases the rate of cAMP production inside the cell.
3-6. Protein Switches Based on Nucleotide Hydrolysis Most protein switches are enzymes that catalyze the hydrolysis of a nucleoside triphosphate to the diphosphate - GTPase : major class of switch protein (G protein) - ATPase : usually associated with motor protein complexes or transporters ATPase GTPase -two-component response regulator: histidine kinase, response regulator proteins Why ATP or GTP are used for trigger of switch? Figure3-12. Structure of the core domains of a typical GTPase and an ATPase
3-6. Protein Switches Based on Nucleotide Hydrolysis • - Triphosphate-bound state = “on”, spring-loaded • Loss of gamma phosphate group → conformational change. • - Two hydrogen bonds in the each switch (Ⅰ and Ⅱ). a- and b- phosphates are bound to P-loop (GXXXXGKS/T) g-phospate is bound to both switch I and II (DXnT and DXXG respectively) Although common structural and functional features in switch proteins, many insertions of other domains in individual GTPases present various functions. Figure3-13. Schematic diagram of the universal switch mechanism of GTPases
3-7. GTPase Switches : Small Signaling G Proteins The switching cycle of nucleotide hydrolysis and exchange in G proteins is modulated by the binding of other proteins GTP hydrolysis rate is very low → GAP(GTPase-activating protein) increase the rate by 105 fold GDP release is conducted by GEF(guanidine-nucleotide exchange factors) Opening up the binding site Figure3-14. The switching cycle of the GTPase involves interactions with proteins that facilitate binding of GTP and stimulation of GTPase activity
Small GTPase Ras family: H-, N-,and K-ras, 21kDa, lipid attachment Signal transduction by Ras is dependent on the GTP-bound state. A prolonged on state are found in up to 30% of human tumors. Reduction of GTP hydrolysis is caused by point mutations at 12, 13 or 61 resulting in uncontrolled cell growth and proliferation. Good target for anti-tumor therapy. How the GAP facilitate GTP hydrolysis? - GAP insert an arginine side chain into the nucleotide-binding site of the GTPase. The positive charge on the side chain helps to stabilize the negative charge in the transition state for hydrolysis of the g-phosphate group of GTP
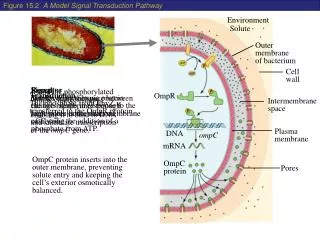
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases Heterotrimeric GTPase α, β and γ subunit. α subunit consist of the canonical G domain and an extra helical domain. β and γ subunit are tightly associated with each other by coiled-coil interaction. G protein associated with G protein coupled receptor(GPCR). GDP-bound G protein bind to GPCR = “off” state. When activated by ligand, these receptors act as GEF for their partner G protein. When GDP is released and GTP binds, G protein dissociates from the GPCR. In the absence of β and γ, α does not bind to GPCR.
Regulator of G-protein signaling proteins (RGS proteins) are responsible for the GTPase catalytic rate. How it increase the rate? a subunit of G-protein has a “built-in” arginine residue in the extra helical domain that projects into the catalytic site. RGS proteins bind to the switch regions, reducing the flexibility and stabilization the transition state for hydrolysis. Particular RGS proteins regulate particular GPCRs; specificity GPCRs are the most numerous receptors in all eukaryotic genome (1-5% of the total number of genes) various ligands such as light, orants, lipids, peptide hormones. 8 families
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases GPCR = “Off” state WD40 Coiled-coil interaction Figure3-15. Hypothetical model of a heterotrimeric G protein in a complex with its G-protein-coupled receptor
3-6. Protein Switches Based on Nucleotide Hydrolysis Most protein switches are enzymes that catalyze the hydrolysis of a nucleoside triphosphate to the diphosphate - GTPase : major class of switch protein (G protein) - ATPase : usually associated with motor protein complexes or transporters ATPase GTPase -two-component response regulator: histidine kinase, response regulator proteins Why ATP or GTP are used for trigger of switch? Figure3-12. Structure of the core domains of a typical GTPase and an ATPase
3-6. Protein Switches Based on Nucleotide Hydrolysis • - Triphosphate-bound state = “on”, spring-loaded • Loss of gamma phosphate group → conformational change. • - Two hydrogen bonds in the each switch (Ⅰ and Ⅱ). a- and b- phosphates are bound to P-loop (GXXXXGKS/T) g-phospate is bound to both switch I and II (DXnT and DXXG respectively) Although common structural and functional features in switch proteins, many insertions of other domains in individual GTPases present various functions. Figure3-13. Schematic diagram of the universal switch mechanism of GTPases
3-7. GTPase Switches : Small Signaling G Proteins The switching cycle of nucleotide hydrolysis and exchange in G proteins is modulated by the binding of other proteins GTP hydrolysis rate is very low → GAP(GTPase-activating protein) increase the rate by 105 fold GDP release is conducted by GEF(guanidine-nucleotide exchange factors) Opening up the binding site Figure3-14. The switching cycle of the GTPase involves interactions with proteins that facilitate binding of GTP and stimulation of GTPase activity
Small GTPase Ras family: H-, N-,and K-ras, 21kDa, lipid attachment • Signal transduction by Ras is dependent on the GTP-bound state. A prolonged on state are found in up to 30% of human tumors. Reduction of GTP hydrolysis is caused by point mutations at 12, 13 or 61 resulting in uncontrolled cell growth and proliferation. Good target for anti-tumor therapy. • How the GAP facilitate GTP hydrolysis? - GAP insert an arginine side chain into the nucleotide-binding site of the GTPase. The positive charge on the side chain helps to stabilize the negative charge in the transition state for hydrolysis of the g-phosphate group of GTP
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases • Heterotrimeric GTPase • α, β and γ subunit. • α subunit consist of the canonical G domain and an extra helical domain. • β and γ subunit are tightly associated with each other by coiled-coil interaction. • G protein associated with G protein coupled receptor(GPCR). • GDP-bound G protein bind to GPCR = “off” state. • When activated by ligand, these receptors act as GEF for their partner G protein. • When GDP is released and GTP binds, G protein dissociates from the GPCR. • In the absence of β and γ, α does not bind to GPCR.
Regulator of G-protein signaling proteins (RGS proteins) are responsible for the GTPase catalytic rate. How it increase the rate? • a subunit of G-protein has a “built-in” arginine residue in the extra helical domain that projects into the catalytic site. RGS proteins bind to the switch regions, reducing the flexibility and stabilization the transition state for hydrolysis. • Particular RGS proteins regulate particular GPCRs; specificity • GPCRs are the most numerous receptors in all eukaryotic genome (1-5% of the total number of genes) • various ligands such as light, orants, lipids, peptide hormones. • 8 families
3-8. GTPase Switches : Signal Relay by Heterotrimeric GTPases GPCR = “Off” state WD40 Coiled-coil interaction Figure3-15. Hypothetical model of a heterotrimeric G protein in a complex with its G-protein-coupled receptor
Cyclic AMP stimulates the phosphorylation of many target proteins by activating protein kinase A • The increased concentration of cAMP → affect a wide range of cellular processes(ATP production stimulation in muscle cell, enhances the degradation of storage fuels in other cell and so on…). • The key enzyme of effects of cAMP is protein kinase A(PKA). • PKA : stimulates the expression of specific genes by phosphorylating a transcriptional activator.
G proteins spontaneously reset themselves through GTP hydrolysis • How is the signal switched off? • Gα subunits have intrinsic GTPase activity(GTP → GDP+Pi). • (The hydrolysis is slow) • - GDP bound αsubunit re-associates with βγsubunits to re-form the inactive heterotrimeric protein.
The hormone bound activated receptor must be reset. • Hormone dissociates. • Remaining receptor-hormone complex is deactivated by the phosphorylation of Ser and Thrin the C’ terminal. • β-Arrestin bind to phosphorylated C-terminal tail and further diminishes its ability to activate G protein.
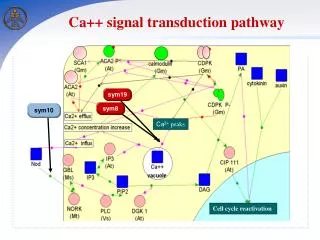
Some 7TM receptors activate the phosphoinositide cascade • Angiotensin II receptor: control of blood pressure • Angiotensin binds to Angiotensin II receptor and activate Gaq • Activated G-protein activates “phospholipase C” • Phsopholipase C catalyzes the cleavage of PIP2 → IP3+DAG.
Free IP3 binds to Ca2+ channel Ca2+ flow out from ER to cytoplasm Ca2+ bind to protein kinase C Ca2+ bind to calmodulin Smooth muscle contraction Glycogen break down Vesicle release…… - DAG remains in the plasma membrane. It activates protein kinase C.
Calcium ion is a widely used second messenger • Why Ca2+ ? • Intracellular low concentration in 100nM • Ca2+ can bind tightly to proteins and induce conformational changes. • Bind to negatively charged oxygen atoms(Glu and Asp). • Coordinated by 6~8 oxygen atoms.
- Changes in Ca2+ concentrations can be monitored in real time. • Fura-2 : specially designed dye. • binds Ca2+ and change their fluorescent properties. • binds Ca2+ through positioned oxygen atoms(red).
When a dye is introduced into cells, changes in available Ca2+ concentration can be monitored with microscopes that detect changes in fluorescence. • Red : high Ca2+ concentration • Blue : low Ca2+ concentration
Calcium ion often activates the regulatory protein calmodulin • ※Calmodulin : 17kda, 4 Ca2+ binding sites. Sensor of Ca2+. Activated by the binding of Ca2+. EF-hand protein family member. • EF-hand protein : Ca2+ binding motif. Two helices(E and F) are positioned like the forefinger and thumb of the right hand.
The binding of to calmodulin induces conformational changes(=expose hydrophobic surface that can be used to bind other proteins)
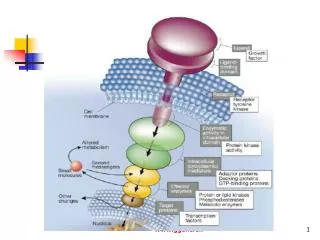
14.2 Insulin signaling : phosphorylation cascades are central to many signal-transduction processes The insulin receptor is a dimer that closes around a bound insulin molecule • Insulin : peptide hormone, 2 chains, linked by 3 disulfide bonds. • - Insulin receptor: dimer of 2 identical units, Each unit consists of 1 αchain and 1 βchain. • (αchain = outside of the cell, ligand binding site. • βchain = inside of the cell, membrane spanning + protein kinase domain)
Insulin binding results in the cross-phosphorylation and activation of the insulin receptor • Insulin binding → 2 α subunits close together • → 2 βsubunits close together • → tyrosine kinase activation When these tyrosine residues phosphorylated, a striking conformational change take place. Insulin binding → tyrosine kinase activation
The activated insulin receptor kinase initiates a kinase cascade - Additional sites within the receptor also are phosphorylated → act as docking site for other substrate(ex>IRS-1). - IRS(insulin receptor substrate) : signal transduction through a series of membrane anchored molecules to a protein kinase.
N-terminal of IRS : pleckstrin homology domain(binds phosphoinositide), phosphotyrosine binding domain. • Center~C-terminal of IRS : 4 YXXM seq. (phosphorylated by insulin receptor tyrosine kinase) - IRS = adaptor proteins ; they bind to the lipid kinase and bring it to the membrane so that it can act on its substrate.
- Phosphotyrosine in the IRS are recognized by SH2 domain. • ※ SH2(Src homology) • Present in many signal transduction proteins. • Bind to phosphotyrosine containing peptide. • Ex> phosphatidylinositide 3- kinase
Contains SH2 domain PIP3 activates PDK1(protein kinase) PDK1 phosphorylates and activates other kinase Akt. Akt phosphorylates targets(ex> GLUT1-glucose transporter)
- The signal is amplified at several stages along this pathway.
Insulin signaling is terminated by the action of phosphatases • Specific enzymes are required to hydrolyze these phosphorylated proteins and convert them back into the states that they were in before the initiation of signaling. • Lipid phosphatase. • Tyrosine phosphatase. • Serine phosphatase……
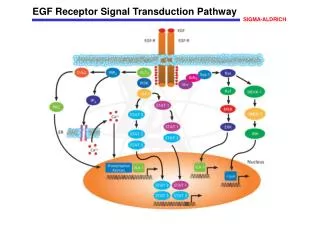
14.3 EGF signaling : signal-transduction pathways are poised to respond EGF binding results in the dimerization of the RGF receptor • ※ EGF • Epidermal growth factor. • 6kda polypeptide.(53 a.a) • Stimulates the growth of epidermal and epithelial cells. • Three intrachain disulfide bonds stabilize the structure.
※ EGF receptor • Protein tyrosine kinase. • Cross-phosphorylation reaction. • Single transmembrane spanning helix. • Dimer of two identical units. Exist as monomers until EGF ligand binding them.
The receptor dimer binds two ligand molecules. • EGF molecule lies far away from the dimer interface. • This interface includes a dimerization arm.
Dimer Monomer • Each monomer is in a conformation that is quite different from that observed in the ligand-bound dimer. • The dimerization arm binds to a domain within the same monomer. (= tethered)
Her2(one of EGF receptor) exists in the extended conformation even in the absence of bound ligand. • 50% identical in amino acid seq. with the EGF receptor and has the same domain structure. • Does not bind any known ligand. • Forms heterodimers with the EGF receptor and other members of the EGF receptor family. • Overexpressed in some cancers. • Overexpression = tumor growth.
The EGF receptor undergoes phosphorylation of its carboxyl-terminal tail • Dimerization → tyrosine kinase activation → C terminal of kinase domain is phosphorylated. • 5 tyrosine residues. • The dimerization of the EGF receptor brings the C-terminal region on one receptor into the active site of its partner’s kinase.
EGF signaling leads to the activation of Ras, a small G protein • The phosphotyrosines on the receptors act as docking sites for SH2 domains on other partner. • Grb-2 : adapter protein. Contains SH2 domain. • recognizes phosphotyrosine residues of EGFR. • then recruits Sos protein through SH3 domain. - Sos binds to Ras(signal transduction component).