Download

1 / 61
610 likes | 727 Views
Signal Transduction T.H. Chiu, September, 2001 I. Introduction II. Signals can enter cells in several ways III. Four general schemes of cell-to-cell signaling IV. Most signals are under complex control of endocrine and nervous systems

E N D
Signal Transduction T.H. Chiu, September, 2001 I. Introduction II. Signals can enter cells in several ways III. Four general schemes of cell-to-cell signaling IV. Most signals are under complex control of endocrine and nervous systems V. Enzymatic cascades result in amplification of extracellular signal VI. Down regulation of cell surface receptors
VII. The same signal molecule can cause different cell responses VIII. Ligand-gated ion channels IX. Four types of “second messengers” are produced by G-protein coupled receptors X. Receptor tyrosine kinases XI. Mechanism of action of steroid hormones
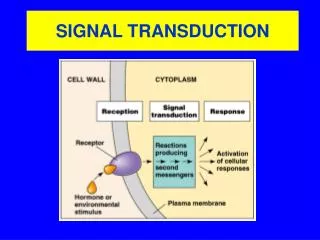
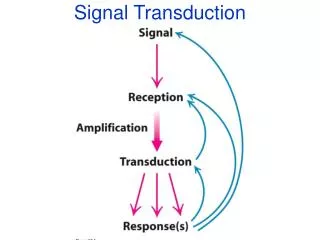
I. Introduction: Signal transduction refers to the process by which external signals (one form of information), such as hormones, neurotransmitters, and growth factors, are transduced into cellular changes (another form of information). Signal transduction plays vital roles in the survival and the death of the cells, including the development, the growth, the maturation, and the aging of the organism. Hence, signal transduction can be considered as the most basic and fundamental activity of the cell.
II. Signal can enter cells in several ways: 1. Steroid receptors: a. passive diffusion of hormone across plasma membrane b. hormone binds cytosolic or nuclear receptors c. receptor acts as enhancer of gene transcription d. cellular response results in new protein and enzyme activities 2. Ligand-gated ion channels: a. neurotransmitters (ACh, GABA) bind to extracellular domain of transmembrane receptor b. open channel allows entry of cations (Na+ and K+ for ACh) or anions (Cl- for GABA)
II. Signals can enter cells in several ways: 3. G-protein receptors: a. variety of ligands bind transmembrane receptors b. receptors coupled to diverse effector enzymes via G-protein c. effector enzymes produce “second messenger” molecules (cAMP, cGMP, phosphoinositides, Ca++)
II. Signals can enter cells in several ways: 4. Transmembrane enzymes a. peptide hormones (insulin, EGF) bind to extracellular domain of transmembrane receptor b. ligand activates tyrosine kinase activity of cytoplasmic domain c. protein phosphorylation on tyrosine residues of tyrosine kinase d. a variety of substrates involved in cellular responses such as ras, raf, MAP kinase pathway but the details remain to be established e. Cardiac glycosides bind to Na+,K+-ATPase, resulting in an increase of intracellular Ca++.
III. Four general schemes of cell-to-cell signaling: 1. Endocrine signaling: Hormone secreted by endocrine organ is carried in blood (usually bound to carrier protein) to distant target cell 2. Paracrine signaling: Signal molecules from secretory cell only affects adjacent cell, they act as local mediators. They may regulate inflammation at sites of infection or cell proliferation in wound healing. A related version of paracrine signaling is autocrine signaling, which refers to cells respond to substance that themselves release. Example: release of growth factors by tumor cells that stimulate unregulated growth of tumor itself as well as adjacent cells
III. Four general schemes of cell-to-cell signaling 3. Neuronal signaling: message carried over long distance but with high specificity as dictated by the transmitters released and receptors activated 4. Contact-dependent signaling: It dose not require the release of a secreted molecules. Instead, the cells make direct contact through signal molecules in their plasma membranes. Example: differentiation of unspecified epithelial cells into nerve cells. Delta molecules located on the plasma membrane of developing nerve cells inhibit the Notch receptors on the epithelial cells, resulting in the inhibition of differentiation (see Fig. 15-4).
IV. Most signals are under complex control of endocrine and nervous systems Example: Thyroid hormone secretion and target cell response 1. Stimulation of brain nerve cells by environmental cues (cold, stress) 2. Nerves act on hypothalamus via neurotransmitters and ligand-gated channels to cause secretion of peptide hormone, TRH (thyrotropin-releasing hormone) 3. TRH acts on pituitary via G-protein receptors (phosphoinositide pathway) to cause secretion of peptide hormone, TSH (thyroid-stimulating hormone)
IV. Most signals are under complex control 4. TSH acts on thyroid gland via G-protein receptors (cAMP pathway) to cause secretion of thyroid hormone (steroid-like molecule) 5. Thyroid hormone acts on most cells via intracellular receptor and gene transcription to increase basal metabolism 6. This relationship is depicted in the following figure
Thyroid hormone secretion and target cell response involve a cascade of complex control
V. Enzymatic cascades result in amplification of extracellular signals An example is shown in the following figure which demonstrates the activation of receptor/G-protein coupled adenylyl cyclase resulting in the formation of cAMP. 1. A ligand concentration of 10-9 M can result in 10-6 M cAMP
VI. Down regulation of cell surface receptors Down regulation of receptors is a general response to high levels of circulating hormone. There are two major ways in which cell surface hormone receptors are “inactivated” in response to hormone: 1. Ligand-induced endocytosis a. common mechanism to insulin, glucagon and EGF receptors b. short-term hormone exposure results in hormone- receptor internalization. Endocytic vesicle and lysosome fuse, resulting in preferential degradation of the peptide hormone. Majority of receptor is recycled to plasma membrane via exocytosis.
VI. Down regulation of cell surface receptors 1. Ligand-induced endocytosis c. Prolonged exposure to hormone results in degradation of both receptor and peptide hormone, markedly reducing receptor numbers at cell surface. d. After lowering of extracellular hormone levels, 12-24 hours of new receptor synthesis are required to normalize receptor numbers. e. An example is shown in the following figure.
Common mechanism to insulin, glucagon and epidermal growth factor receptors Short-term hormone exposure results in hormone-receptor internalization. Endocytic vesicle and lysosome fuse, resulting in preferential degradation of the peptide hormone. Majority of receptor is recycled to plasma membrane via exocytosis. Prolonged exposure results in degradation of both receptor and peptide hormone, markedly reducing receptor numbers at cell surface. After lowering of extracellular hormone levels, 12-24 hours of new receptor synthesis are required to normalize receptor numbers.
VI. Down regulation of cell surface receptors 2. Desensitization via phosphorylation: an example is given below for β-adrenoceptor a. Prolonged exposure (hrs) results in β-adrenoceptors which can still bind hormone but which cannot activate adenylyl cyclase b. Desensitization occurs thru phosphorylation of serine residue on the cytoplasmic domain of the receptors c. Two kinases are involved: i. β-adrenoceptor kinase (βARK) phosphorylates only the agonist-bound receptor ii. cAMP-dependent protein kinase (A-kinase) phosphorylates and inactivates the receptor when intracellular cAMP levels get too high
VII. The same signal molecule can cause different cell responses ACh causes contraction of skeletal muscle, but relaxation of heart muscle. Due to different receptors in each cell type. Nicotinic ACh receptors of skeletal muscle are ligand-gated channels, while muscarinic ACh receptors of heart muscle are of the G-protein type. Both heart muscle and secretory cells (e.g., pancreatic cells) contain identical ACh receptors (G-protein), but signal induces relaxation in one and secretion of insulin in the other. Differential response due to cell-type specific effector moleules.
VIII. Ligand-gated ion channels They convert extracellular signal into electrical impulse. They are located at specialized junctions between nerve cells and target cells (chemical synapses) 1. Nerve terminal releases neurotransmitters (ACh, GABA, excitatory amino acids) by fusion of storage vesicles with plasma membrane 2. Neurotransmitter binds ligand-gated plasma membrane receptors concentrated in synapse region of target cell 3. Gated channel opens allowing entry of ions into target cell
Nicotinic acetylcholine receptor 1. Channel opening requires 2 molecules of ACh 2. Channel remains open only briefly (~ 1 millisecond) and closes even though ligand is still bound 3. ACh dissociates from receptor returning it to resting state 4. Dissociated ACh is hydrolyzed by acetylcholinesterase located at neuromuscular junction. Choline is actively taken up by the nerve terminal for the resynthesis of ACh.
Structure of nicotinic ACh receptor 1. Composed of 5 transmembrane polypeptides: two α and one each for β,γ, and δ. 2. α chains contain ligand-binding sites. Each chain traverses plasma membrane 4 times 3. One helix of α chain contains more polar amino acids, forming wall of “aqueous pore” (hydrophilic channel) 4. Clusters of negatively charged amino acids at the mouth of the channel serve to prevent entry of negative ions 5. Any cation with diameter of less than 0.65 nanometers can pass through, usually Na+ and K+ but some Ca++
IX. Four types of “second messengers” are produced by G-protein coupled receptors 1. cyclic AMP (cAMP) 2. diacyglycerol (DAG) 3. Ca++/phosphoinositides 4. cyclic GMP (cGMP) Major hormone responses mediated by cAMP: thyroid stimulating hormone, ACTH, LH, epinephrine, parahormone, vasopressin, glucagon Depending on cell type, the same hormone (epinephrine) and the same second messenger (cAMP) can result in very different cellular responses
Structure of the β-adrenoceptor 1. single polypeptide chain which traverses the plasma membrane 7 times 2. extracellular amino terminus contains ligand-binding site; intracellular carboxy terminus contains sites for G- protein interaction and phosphorylation-mediated desensitization 3. most G-protein receptors are 7-pass transmembrane proteins Examples: muscarinic ACh receptor α-adrenoceptor rhodopsin
Model of receptor coupling to adenylyl cyclase via stimulatory G-protein: 1. Three major components: a. receptor b. multimeric G-protein c. adenylyl cyclase 2. Receptor and adenylyl cyclase are transmembrane proteins, but G-protein is located only at cytoplasmic face of plasma membrane 3. Gs is composed of three polypeptide chains: αs chain - binds and hydrolyzes GTP, interacts with receptor, and activates adenylyl cyclase β, γ chains - form tight complex, anchors αs chain to cytoplasmic face of membrane, both chains identical for stimulatory and inhibitory alpha subunits
Synthesis and degradation of cyclic AMP 1. resting concentration of cAMP = 10-6 M 2. hormonal stimulation results in rapid (seconds) increase (5-fold) in cAMP oncentration 3. responsiveness requires rapid synthesis of cAMP balanced by rapid breakdown 4. adenylyl cyclase synthesizes cAMP from ATP by cleaving off pyrophosphate 5. cAMP phosphodiesterase hydrolyzes cAMP to 5’-AMP 6. synthesis of cAMP from ATP is irreversible due to pyrophosphatase which will hydrolyze pyrophosphate released by adenylyl cyclase
Model of cAMP-mediated cellular response 1. cAMP activates cAMP-dependent protein kinases (R2C2) 2. kinases are tetramers: two (2) cAMP-binding regulatory (R) chains two (2) catalytic (C) chains 3. binding of cAMP causes release of activated catalytic units 4. catalytic units phosphorylate a variety of substrates, usually enzymes, which are either activated or inactivated 5. enzymatic activity or lack there of then induces the final cellular response to hormone 6. Moderation and reversal f cAMP response is achieved via (a) dephosphorylation of substrates by phosphatases and (b) degradation of cAMP to 5’-AMP by PDE
Example of cAMP-mediated cellular response: Epinephrine-induced breakdown of liver and muscle glycogen 1. Epinephrine binds β-adrenoceptor and initiates production of cAMP via G-protein coupled adenylyl cyclase 2. cAMP activates cAMP-dependent protein kinase (A-kinase) 3. A-kinase phosphorylates and activates phosphorylase kinase 4. Phosphorylase kinase phosphorylates and activates glycogen phosphorylase, the enzyme that breaks down glycogen 5. A-kinase also phosphorylates and inactivates the enzyme responsible for glycogen synthesis, glycogen synthase
Adenylyl cyclase can be inactivated by inhibitory G-protein Same signal molecule can either increase or decrease intracellular cAMP concentrations depending on the type of receptor bound. Other ligands will only decrease cAMP concentration. In either case, the ligand causes cAMP reduction by binding to a receptor that is linked to an “inhibitory” G-protein, Gi. 1. Gi is composed of: a. β and γ subunits identical to those found in Gs. b. a distinct αi subunit which can bind and hydrolyze GTP but which inactivates adenylyl cyclase by unknown mechanism 2. Examples of ligands and receptors which lower cAMP via Gi: α2-adrenoceptor; PGE1 and adenosine
The cGMP signaling pathway Unlike cAMP, cGMP plays a significant role in only a few organ systems: 1. Photoreceptor of the eye: A protein-linked mechanism involving a 7-pass trans-membrane receptor (rhodopsin), a unque G protein (tranducin), and cGMP phosphodiesterase (PDE) 2. Vascular smooth muscle a. Atrial natriuretic peptide (ANP) produced in heart, regulates blood volume and vascular tone via trans-membrane receptor with intrinsic guanylyl cyclase activity b. Nitric oxide (NO), produced in vascular endothelial cells in response to natural vasodilator agents such as ACh and histamine, acts on vascular smooth muscle via cytoplasmic guanylyl cyclase
cGMP signaling pathway in the visual system PDE: phosphodiesterase
Amplification in the light-induced signaling cascade in rod photoreceptor cell
The Ca++/phosphoinositide signal pathway Even more complicated than the cAMP pathway, as three messenger molecules are produced: 1. diacylglycerol 2. inositol triphosphate (IP3) 3. Ca++ Major cellular responses utilizing this system are: liver/vasopressin/glycogen breakdown pancreas/ACh/amylase secretion smooth muscle/ACh/contraction pancreatic βcells/ACh/insulin secretion mast cells/antigen/histamine release blood platelets/thrombin/serotonin and PDGF secretion - both organic (ACh) and peptide (e.g., thrombin) hormones activate this pathway - ACh receptor is of the “muscarinic” type