Download

1 / 16
160 likes | 280 Views
Cell to Cell Communication Yasir Waheed. Introduction.

E N D
Cell to Cell Communication Yasir Waheed
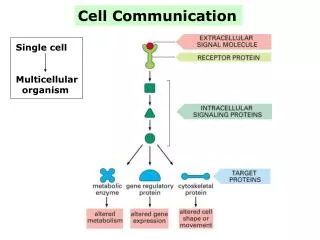
Introduction • Present-day bacteria were present on Earth for about 2.5 billion years before the first multicellular organisms appeared. One reason why multicellularity was so slow to evolve may have been related to the difficulty of developing the elaborate cell communication mechanisms that a multicellular organism needs. • These communication mechanisms depend heavily on extracellular signal molecules, which are produced by cells to signal to their neighbors or to cells further away. This system involves elaborate systems of proteins include cell-surface receptor proteins, which bind the signal molecule, plus a variety of intracellular signaling proteins that distribute the signal to appropriate parts of the cell. Among the intracellular signaling proteins are kinases, phosphatases, GTP-binding proteins, and many other proteins with which they interact. At the end of each intracellular signaling pathway are target proteins, which are altered when the pathway is active and change the behavior of the cell.
Figure 15-1. A simple intracellular signaling pathway activated by an extracellular signal molecule. The signal molecule binds to a receptor protein (which is usually embedded in the plasma membrane), thereby activating an intracellular signaling pathway that is mediated by a series of signaling proteins. Finally, one or more of these intracellular signaling proteins interacts with a target protein, altering the target protein so that it helps to change the behavior of the cell.
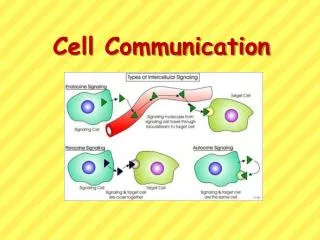
Forms of intercellular signaling. Figure 15-4. Forms of intercellular signaling. (A) Contact-dependent signaling requires cells to be in direct membrane-membrane contact. (B) Paracrine signaling depends on signals that are released into the extracellular space and act locally on neighboring cells. (C) Synaptic signaling is performed by neurons that transmit signals electrically along their axons and release neurotransmitters at synapses, which are often located far away from the cell body. (D) Endocrine signaling depends on endocrine cells, which secrete hormones into the bloodstream that are then distributed widely throughout the body.
Each Cell Is Programmed to Respond to Specific Combinations of Extracellular Signal Molecules Figure 15-8. An animal cell's dependence on multiple extracellular signals. Each cell type displays a set of receptors that enables it to respond to a corresponding set of signal molecules produced by other cells. These signal molecules work in combinations to regulate the behavior of the cell. As shown here, an individual cell requires multiple signals to survive (blue arrows) and additional signals to divide (red arrow) or differentiate (green arrows). If deprived of appropriate survival signals, a cell will undergo a form of cell suicide known as programmed cell death, or apoptosis.
Ligand may bind to intracellular or extracellular receptor Figure 15-3. The binding of extracellular signal molecules to either cellsurface receptors or intracellular receptors. Most signal molecules are hydrophilic and are therefore unable to cross the plasma membrane directly; instead, they bind to cell-surface receptors, which in turn generate one or more signals inside the target cell. Some small signal molecules, by contrast, diffuse across the plasma membrane and bind to receptors inside the target cell either in the cytosol or in the nucleus (as shown here). Many of these small signal molecules are hydrophobic and nearly insoluble in aqueous solutions; they are therefore transported in the bloodstream and other extracellular fluids after binding to carrier proteins, from which they dissociate before entering the target cell.
Different Cells Can Respond Differently to the Same Extracellular Signal Molecule Figure 15-9. Various responses induced by the neurotransmitter acetylcholine. Different cell types are specialized to respond to acetylcholine in different ways. (A and B) For these two cell types, acetylcholine binds to similar receptor proteins, but the intracellular signals produced are interpreted differently in cells specialized for different functions. (C) This muscle cell produces a distinct type of receptor protein for acetylcholine, which generates different intracellular signals from the receptor shown in (A) and (B), and results in a different effect. (D) The chemical structure of acetylcholine.
The role of nitric oxide (NO) in smooth muscles. Figure 15-11. The role of nitric oxide (NO) in smooth muscle relaxation in a blood vessel wall. Acetylcholine released by nerve terminals in the blood vessel wall activates NO synthase in endothelial cells lining the blood vessel, causing the endothelial cells to produce NO. The NO diffuses out of the endothelial cells and into the underlying smooth muscle cells, where it binds to and activates guanylyl cyclase to produce cyclic GMP. The cyclic GMP triggers a response that causes the smooth muscle cells to relax, enhancing blood flow through the blood vessel.
Figure 15-13. The nuclear receptor superfamily. All nuclear hormone receptors bind to DNA as either homodimers or heterodimers, but for simplicity we show them as monomers here. (B) A receptor protein in its inactive state is bound to inhibitory proteins. Domain-swap experiments suggest that many of the ligand-binding, transcription-activating, and DNA-binding domains in these receptors can function as interchangeable modules. (C) The binding of ligand to the receptor causes the ligand-binding domain of the receptor to clamp shut around the ligand, the inhibitory proteins to dissociate, and coactivator proteins to bind to the receptor's transcription-activating domain, thereby increasing gene transcription.
Figure 15-14. Responses induced by the activation of a nuclear hormone receptor. (A) Early primary response and (B) delayed secondary response. The figure shows the responses to a steroid hormone, but the same principles apply for all ligands that activate this family of receptor proteins. Some of the primary-response proteins turn on secondary-response genes, whereas others turn off the primary-response genes. The actual number of primary- and secondary-response genes is greater than shown. As expected, drugs that inhibit protein synthesis suppress the transcription of secondary-response genes but not primary response genes.
Figure 15-16. Different kinds of intracellular signaling proteins along a signaling pathway from a cell-surface receptor to the nucleus.
Relay proteins simply pass the message to the next signaling component in the chain. • Messenger proteins carry the signal from one part of the cell to another, such as from the cytosol to the nucleus. • Adaptor proteins link one signaling protein to another, without themselves conveying a signal. • Amplifier proteins, which are usually either enzymes or ion channels, greatly increase the signal they receive, either by producing large amounts of small intracellular mediators or by activating large numbers of downstream intracellular signaling proteins. When there are multiple amplification steps in a relay chain, the chain is often referred to as a signaling cascade. • Transducer proteins convert the signal into a different form. The enzyme that makes cyclic AMP is an example: it both converts the signal and amplifies it, thus acting as both a transducer and an amplifier. • Bifurcation proteins spread the signal from one signaling pathway to another. • Integrator proteins receive signals from two or more signaling pathways and integrate them before relaying a signal onward. • Latent gene regulatory proteins are activated at the cell surface by activated receptors and then migrate to the nucleus to stimulate gene transcription. • Modulator proteins modify the activity of intracellular signaling proteins and thereby regulate the strength of signaling along the pathway. • Anchoring proteins maintain specific signaling proteins at a precise location in the cell by tethering them to a membrane or the cytoskeleton. • Scaffold proteins are adaptor and/or anchoring proteins that bind multiple signaling proteins together in a functional complex and often hold them at a specific location.
Some Intracellular Signaling Proteins Act as Molecular Switches Figure 15-17. Two types of intracellular signaling proteins that act as molecular switches. In both cases, a signaling protein is activated by the addition of a phosphate group and inactivated by the removal of the phosphate. (A) The phosphate is added covalently to the signaling protein by a protein kinase. (B) A signaling protein is induced to exchange its bound GDP for GTP. To emphasize the similarity in the two mechanisms, ATP is shown as APPP, ADP as APP, GTP as GPPP, and GDP as GPP.
Figure 15-18. Signal integration. (A) Extracellular signals A and B both activate a different series of protein phosphorylations, each of which leads to the phosphorylation of protein Y but at different sites on the protein. Protein Y is activated only when both of these sites are phosphorylated, and therefore it becomes active only when signals A and B are simultaneously present. For this reason, integrator proteins are sometimes called coincidence detectors. (B) Extracellular signals A and B lead to the phosphorylation of two proteins, a and b, which then bind to each other to create the active protein. In both of the examples illustrated, the proteins themselves are phosphorylated.
Figure 15-25. Five ways in which target cells can become desensitized to a signal molecule. The inactivation mechanisms shown here for both the receptor and the intracellular signaling protein often involve phosphorylation of the protein that is inactivated, although other types of modification are also known to occur. In bacterial chemotaxis, which we discuss later, desensitization depends on methylation of the receptor protein.