Download
1 / 32
440 likes | 1.22k Views
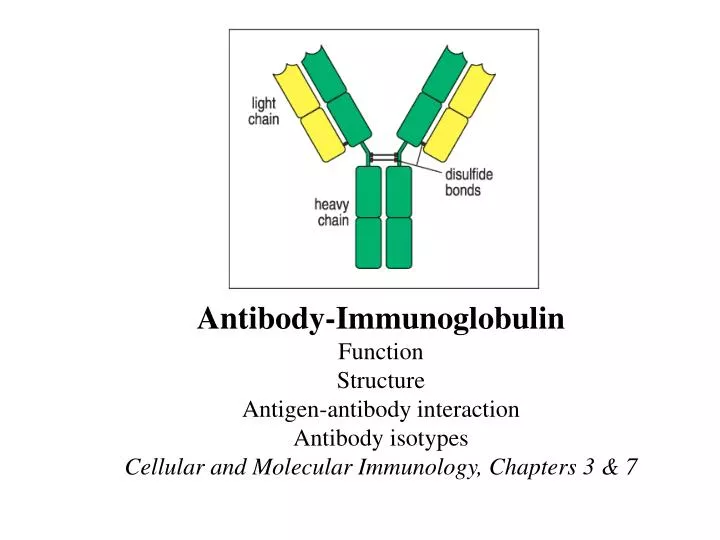
Antibody-Immunoglobulin Function Structure Antigen-antibody interaction Antibody isotypes Cellular and Molecular Immunology, Chapters 3 & 7. Web. Structures of antibodies. Two heavy chains and two light chains Disulfide bonds Ig domains Hinge region Constant and variable regions

E N D
Antibody-Immunoglobulin Function Structure Antigen-antibody interaction Antibody isotypes Cellular and Molecular Immunology, Chapters 3 & 7
Structures of antibodies • Two heavy chains and two light chains • Disulfide bonds • Ig domains • Hinge region • Constant and variable regions • Two identical antigen-binding sites • Two function domains • a. Bind to antigens • b. Recruit effector cells and molecules • 7. Heavy chains decide the class
Determining Ab Structure • Work in 1950s and 60s using biochemical techniques • Rodney Porter used partial proteolysis with papain - 2 identical antigen-binding fragments (Fab) and one “tail” fragment (Fc) • Alfred Nisinoff used pepsin - one fragment with divalent antigen binding (F(ab2)’) • Gary Edelman used b-ME to reduce Ig, resolving heavy (H) and light (L) chains • Rodney Porter probed H and L chains with anti- Fab and anti-Fc antibodies: anti-Fab detected both H & L, but anti-Fc detected only H • Combined work earned Porter and Edelman 1972 Nobel Prize
Fab-Fragment antigen binding Fc-Fragment crystallizable Fv-Fragment variable Fig. 3.3 The Y-shaped immunoglobulin molecule can be dissected by partial digestion with proteases.
Fine Structure of Ab • “Variable” (V) and “Constant” (C) regions, defined by degree of sequence similarity between different antibodies • Repeated structural motif (“immunoglobulin fold”): antiparallel beta strands form 2 beta sheets; intrachain disulfide leads to beta barrel structure • V-type fold has 9 beta strands, C-type fold has 7 • L chain has one V and one C domain; H chain has one V and 3-4 C domains • Ag-binding site formed by VH + VL
Structure of Ig Light Chain Showing V- and C-type Immunoglobulin Folds
Variability of V-Region • Variability isn’t evenly distributed: “hyper-variable” (HV) regions surrounded by less variable “framework” regions (FR) • Defined by variability plots (number of different a.a./frequency of most common a.a.) • HV regions correspond to loops between beta strands: B-C, F-G, and C’-C”, and can vary in both sequence and length • HV regions also called “complementarity determining regions (CDR), because they make up the Ag-binding surface (H + L)
HV-Hypervariable region CDR-Complementarity-Determining Regions FR-Framework region FR1 FR2 FR3 FR4
The Immunological Conundrum:How is Variability Generated? One gene One protein 1012 genes ??? 1012 antigens
Generating Diversity: DNA Recombination • In 1976, Hozumi and Tonegawa showed that Ab-producing cells have different DNA sequence than other somatic cells in Ig locus - evidence for DNA recombination (1987 Nobel Prize) • Sequencing of Ig genes has identified gene segments that are brought together by recombination
Ig Locus Rearrangement • Sequence-specific: recombination signal is a conserved heptamer (CACAGTG), a spacer (12 or 23 non-conserved bases), and a conserved nonamer (ACAAAAACC) • 12/23 Rule: Recombination machinery always joins a gene segment with a 12-bp spacer to another with a 23-bp spacer • This ensures that the correct gene segments get joined (no V-V, H chain always has D between V and J, etc.)
V(D)J Recombination Machinery • Uses both specific (RAG1/2) and ubiquitous factors • RAG1/2 complex is the sequence-specific recombinase: recognizes recomb signal, brings a 12 and a 23 signal together, and cleaves DNA • Cleaved DNA is repaired by general DNA repair factors
Heavy – 65 (V); 27 (D); 6 (J) = 10,530 Kappa - 40 (V); 0 ; 5 (J) = 200 320 Lambda - 30 (V) 0 ; 4 (J) = 120 10,530 X 320 = 3 million (about 30 million in human, due to more V segments) Actually much much much more! (Junctional diversity, somatic mutations, nucleotide insertions, etc)
Great Diversity Allows for Exquisite Specificity • Antibodies can distinguish between antigens differing by only small chemical substituents • Precise part of an antigen recognized by the antibody is called an epitope, or determinant • Small chemical epitope = hapten • Linear epitope • Conformational epitope (generated by folding)
Antigenic determinant or epitope: The structure recognized by an antibody Haptens Continuous Discontinuous or or linear epitope conformational epitopes Fig. 3.8 Antigens can bind in pockets or grooves, or on extended surfaces in the binding sites of antibodies.
Functions of antibodies • Antibodies neutralize bacterial toxins and complete virus particles and bacterial cells. • Antibodies coating an antigen render it recognizable as foreign by phagocytes (macrophages and neutrophils), which then ingest and destroy it; this is called opsonization. • Antibodies activate of the complement system by coating a bacterial cell. • Fig. 1.24 Antibodies can participate in host defense in three main ways.
Antibody Functions • Ab’s have two “business” ends: antigen-binding arms and Fc tail. • Effector function is mediated by Fc region • Different classes (isotypes) use distinct CH gene segments, and have different effector functions • Isotype switching occurs by another DNA recombination event (except IgM/IgD, which uses alternative RNA splicing)
Isotypes of Immunoglobulin Fig. 4.17 The structural organization of the main human immunoglobulin isotype monomers.
Monoclonal Antibodies • Monoclonal Antibodies - identical antibodies produced in large quantities by an immortalized hybridoma cell line • Technique developed by Köhler and Milstein in 1975, earned Nobel Prize in 1984 • Benefits: identical antibodies available in unlimited quantity; easily purified • Drawbacks: may not be useful for all techniques; limited number of species (mouse, rat, hamster, rabbit)
Production of monoclonal antibodies. HGPRT-hypoxanthine:guanine phosphoribosyl transferase PEG-polyethylene glycol HAT-hypoxanthine-aminopterin-thymidine Cells lacking HGPRT are sensitive to the HAT medium Unfused B cells die within 1-2 days without stimulation