Download

1 / 66
740 likes | 1.43k Views
Antibody structure and function. Parham – Chapter 2. Outline. Antibody structure Antigens Antigen-antibody interactions Generation of antibody diversity Isotype switching Applications - immunoassays. Immunoglobulins – membrane-bound and soluble receptors.

E N D
Antibody structure and function Parham – Chapter 2 H. HogenEsch, 2005
Outline • Antibody structure • Antigens • Antigen-antibody interactions • Generation of antibody diversity • Isotype switching • Applications - immunoassays H. HogenEsch, 2005
Immunoglobulins – membrane-bound and soluble receptors H. HogenEsch, 2005
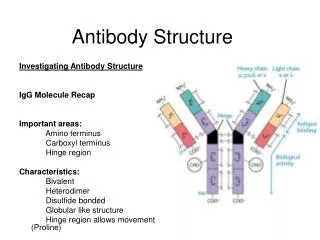
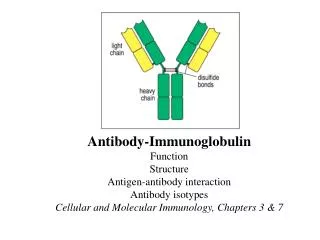
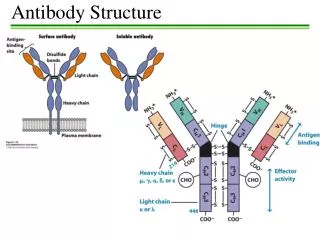
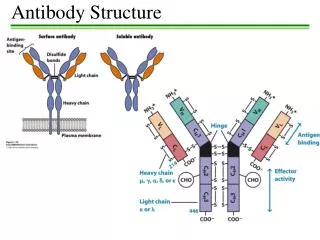
Basic structure of immunoglobulins Fig. 2.2 H. HogenEsch, 2005
Basic structure of immunoglobulins Fig. 2.2 Fig. 2.2 H. HogenEsch, 2005
Antigen-binding Fragment Crystallizable Fragment H. HogenEsch, 2005
g a m d e H: Immunoglobulin classes (isotypes) L-chain: k or l H. HogenEsch, 2005
Structure of immunoglobulins H. HogenEsch, 2005
Structure of immunoglobulins H. HogenEsch, 2005
Hypervariable and framework regions CDR = complementarity -determining region Fig. 2.7 H. HogenEsch, 2005 Fig. 2.7
Differences between immunoglobulins H. HogenEsch, 2005
Epitopes • Epitope (antigenic determinant) is the part of an antigen to which an antibody binds. • Most antigens have multiple epitopes (multivalent) • Usually carbohydrate or peptide. Fig. 2.9 H. HogenEsch, 2005
Immunoglobulin epitopes are usually located at the antigen’s surface. Fig. 2.8 H. HogenEsch, 2005
Conformational vs. linear epitopes Fig. 2.11 H. HogenEsch, 2005
Epitopes heat, acid Conformational epitopes - destroyed by denaturation Linear epitopes - unaffected by denaturation H. HogenEsch, 2005
Epitope recognition H. HogenEsch, 2005
Haptens Small molecules that are not immunogenic by themselves, but can bind immunoglobulins or TCRs. Haptens can induce an immune response when linked to a larger protein. H. HogenEsch, 2005
Hapten Parham Fig. 10.25 H. HogenEsch, 2005
Hapten Parham Fig. 10.26 H. HogenEsch, 2005
Antibody-antigen interaction H. HogenEsch, 2005 Fig. 2.10
Antibody-antigen interaction • Non-covalent binding: • Electrostatic • Hydrogen bonds • Van der Waals forces • Hydrophobic forces • Affinity: Strength of interaction between epitope and one antigen-binding site • Avidity: Strength of the sum of interactions between antibody and antigen Short range H. HogenEsch, 2005
Crossreactivity Antiserum raised against antigen A reacts also with antigen B Antigen A and B share epitopes Antigen A and B have similar (but not identical) epitopes H. HogenEsch, 2005
A B Crossreactivity H. HogenEsch, 2005
Immunoglobulin genes Fig. 2.13 H. HogenEsch, 2005
Somatic recombination – light chain Fig. 2.14 H. HogenEsch, 2005
Somatic recombination – Heavy chain Fig. 2.14 H. HogenEsch, 2005
Number of gene segments Fig. 2.15 H. HogenEsch, 2005
Recombination Signal Sequences Fig. 2.16 H. HogenEsch, 2005
Recombination V(D)J – recombinase Fig. 2.17 H. HogenEsch, 2005
V k J k C k k chain polypeptide V k C k germline DNA // 5’ 3’ 1 2 3 4 5 n 1 2 3 4 5 rearrangement 5’ 3’ B cell DNA V2J3 transcription 5’ 3’ primary RNA transcript splicing mRNA V2J3C translation H. HogenEsch, 2005
Generation of diversity • k chain: 40 V x 5 J = 200 Vk • l chain: 30 V x 4 J = 120 Vl • H chain: 65 V x 27D x 6 J = 10,530 VH • (200 + 120) x 10,530 = 3.4 x 106 combinations H. HogenEsch, 2005
Mechanisms for additional diversity in immunoglobulins • Imprecise joining of gene segments • Random nucleotide addition at joining regions • terminal deoxynucleotidyl transferase (TdT) Fig. 2.17 H. HogenEsch, 2005
Generation of diversity • Multiple gene segments: • - k chain: 40 V x 5 J = 200 Vk • - l chain: 30 V x 4 J = 120 Vl - H chain: 65 V x 27D x 6 J = 10,530 VH • Combination of H and L chain: (200 + 120) x 10,530 = 3.4 x 106 combinations • Imprecise joining and nucleotide addition > 108 different specificities H. HogenEsch, 2005
Organization of CH genes Fig. 2.19 H. HogenEsch, 2005
Naïve mature B cells express IgM and IgD Fig. 2.20 H. HogenEsch, 2005
Allelic exclusion Allelic exclusion ensures that the B lymphocyte expresses immunoglobulin molecules with only one specificity. Mechanism: Successful rearrangement of immunoglobulin gene segmentsone allele shuts down the rearrangement process of the other allele. l k H 16 6 12 H. HogenEsch, 2005
B cell receptor complex Fig. 2.21 H. HogenEsch, 2005
Changes in B cells after activation by antigen • Somatic mutation – additional diversity • Isotype switching H. HogenEsch, 2005
Somatic hypermutation Fig. 2.24 H. HogenEsch, 2005
Hypervariable and framework regions CDR = complementarity -determining region Fig. 2.7 H. HogenEsch, 2005
Isotype switching IgG1 IgG2 IgG3 IgG4 IgM+/IgD+ IgA1 IgA2 IgE H. HogenEsch, 2005
Organization of CH genes Fig. 2.19 H. HogenEsch, 2005
Isotype switching H. HogenEsch, 2005
Physical properties of immunoglobulins H. HogenEsch, 2005
IgM • Membrane-bound monomer and secreted pentamer. • First immunoglobulin to be synthesized during ontogeny and in the immune response. • Activates complement pathway; agglutination. • Can be transported into mucosal secretions. H. HogenEsch, 2005
IgG • Highest concentration in serum. • Four subclasses: IgG1 - 4 • Activates complement • Binds to Fcg -receptors on neutrophils, macrophages and NK cells H. HogenEsch, 2005
IgA • Usually dimer • Secretory IgA is a dimer with a secretory component. • Two subclasses: IgA1 and IgA2 • Major immunoglobulin in mucosal secretions • Neutralization; Prevents binding of micro-organisms to receptors • Not effective activator of complement H. HogenEsch, 2005
IgE • Very low serum concentration in healthy individuals. • Concentration is higher in patients with helminth infections and often in patients with allergies. • Lacks hinge region; extra CH domain • Binds to Fce receptor on mast cells and basophils. Cross-linking results in degranulation and release of pro-inflammatory mediators. H. HogenEsch, 2005
IgD • Very low concentration in serum • Primarily found with IgM on naïve mature B cells • Function is unknown H. HogenEsch, 2005