Download

1 / 44
440 likes | 638 Views
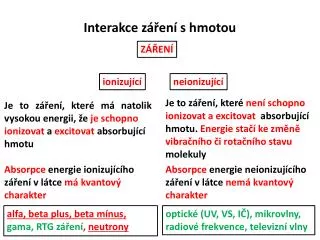
Interakce mezi populacemi. Ovlivnění fitness = působení na natalitu a mortalitu . (popř. růst). Kdybychom vybrali jiné parametry, tak to může dopadnout jinak. - predace - mezidruhová kompetice 0 - komenzalismus 0 - amensalismus - mutualismus. Komenzalismus

E N D
Interakce mezi populacemi
Ovlivnění fitness = působení na natalitu a mortalitu. (popř. růst). Kdybychom vybrali jiné parametry, tak to může dopadnout jinak. -predace -mezidruhová kompetice 0 -komenzalismus 0 -amensalismus -mutualismus
Komenzalismus jeden umožňuje (vylepšuje) existenci druhého. Mohl by sem patřit např. vztah mezi saprofytickými houbami a vyššími rostlinami. Amensalismus typickým příkladem jeallelopatieu rostlin - toxické metabolity jednoho druhu potlačují růst druhu jiného. Mutualismus z evolučního hlediska nejpozoruhodnější, klasičtí ekologové se tím (bohužel) moc nevzrušují. Nicméně ukázkový příklad - a pokud bude čas, ještě se k mutualismu vrátíme
Mezidruhová kompetice Oba zúčastnění ztrácejí fitness. Nicméně to v reálných podmínkách vypadá tak, že jeden je na tom vždy o poznání hůře - kompetice se může jevit víceméně jednostranná (tváří se jako amensalismus). • U kompetice platí: • působí prostřednictvím kombinovaných změnnatalitya mortality • je reciproční • soutěží se o společný zdroj, jehož množství je limitující • je závislá na denzitě Rozdíly v charakteru kompetice mezi pohyblivými a sesilními organismy ( živočichy a rostlinami) Zvířata (pohyblivá, ne sesilní): většina toho, co potřebují k bytí a co může být limitující, je produktem dalších živých organismů - kompetice u zvířat je proto především o potravu. Přitom se málokdy jedná o nepřetržitý boj mezi několika jedinci, častěji se kompetitoři vůbec nepotkají (jiná denní aktivita, jiný způsob vyhledávání potravy apod.) Rostliny: převážně sesilní, a proto v konfliktu s nejbližšími sousedy o místo (živiny v půdě, dopadající světlo).
Mezidruhová kompetice - příklad semenožravých mravenců a hlodavců v pouštích amerického jihozápadu (Brown & Davidson, 1977). 1. Dá se kompetice in natura vůbec pozorovat? 2. Jestli ano, jakou má formu? Obě gildy sdílejí stejný zdroj = velikost semen v potravě se překrývá. Gilda mravenců a gilda hlodavců JE závislá na semenech.
Kompetice existuje! Pokud se jeden z kompetitorů odstranil, četnost druhého vzrostla. Přitom sežral stejně semen jako předtím oba dohormady. Mezidruhová kompetice - příklad semenožravých mravenců a hlodavců v pouštích amerického jihozápadu (Brown & Davidson, 1977). Pokračování pokusu už jen s mravenci: vztah mezi velikostí těla dělníků a velikostí semen, která „sklízeli“. Body jsou průměry a proto neukazují existující překryv. Korelace je velmi zřetelná - větší mravenci žerou větší semena.
Mezidruhová kompetice - příklad semenožravých mravenců v pouštích amerického jihozápadu (Davidson, 1977). Mravenci se liší ve strategii vyhledávání semen. G- hledají ve skupinách, a jen občas. Mají větší úspěšnost, jsou-li semena hojně a ve shlucích. I- individualisti, hledají neustále v celém prostoru. Úspěšnější, pokud se semena vyskytují řídce a rovnoměrně. - na daném místě hojný výskyt - na daném místě vzácný Ve velikostních třídách od 3 mm: pokud jsou na jednom místě stejně velcí, tak se liší v potravní strategii (na lokalitě A výjimka, ale tam je P.m. velmi vzácný Z opačného pohledu: druhy, které jsou stejně velké a mají stejnou strategii, se spolu nevyskytují (nebo je ten druhý velmi vzácný). druhy, které se neliší, spolu nemohou koexistovat.
Mezidruhová kompetice - příklad semenožravých mravenců v pouštích amerického jihozápadu (Davidson, 1978). Pro mravence menší než 3 mm to neplatí. Proč? A) buď toto pravidlo není univerzální, tj. i když sdílí stejný zdroj, tak mohou koexistovat. B) nebo se přeci jen nějak liší, ale nevypátralo se, jak. C) nebo nejsou limitováni semeny, ale něčí jiným, např. predací. Pokračování již jen pro jeden druh - Veromessor pergandei Populace z jednotlivých lokalit se liší, a to nejenom v velikosti, ale hlavně v míře variability. Měřila se délka mandibul. Na lokalitách svyšším počtem kompetitorů menší variabilita.
Mezidruhová kompetice - příklad semenožravých mravenců v pouštích amerického jihozápadu (Davidson, 1978). V. pergandei se na lokalitách s možnou mezidruhovou kompeticí více specializuje. Aktuální velikost mandibul pro danou lokalitu je přitom ovlivněna druhy s nejbližší velikostí mandibul. Nedá se jednoznačně přímo vyvodit, že V. pergandei soutěží s ostatními druhy o semena. Avšak můžeme říci, že na každé lokalitě se adaptoval tak, aby se kompetici co nejvíce vyhnul. Souhrn: 1. Vzdáleně příbuzné taxony mohou soutěžit o limitující zdroj. 2. Kompetice nemusí vést nutně k potlačení jednoho ze soutěžících. 3. Koexistující populace mají tendenci se odlišovat ve způsobu využití limitujícího zdroje. Pokud to nejde, vydrží jen jeden.
Dva sladkovodní plži - Hydrobia ulna a H. ventrosa - pokud je na lokalitě každý sám, tak shodné rozměry a také velikost potravních částic. Pokud pohromadě, tak je velikost ulity odlišná. Druh s větší ulitou preferuje větší potravní částice.
Čmeláci - délka sosáku Na svazích Skalistých hor v Coloradu - Pyke 1982. Čmeláci se dali rozdělit do 4 skupin, podle délky sosáku preferovali i rostliny s adekvátně hlubokým kalichem. (Čmelák ve čtvrté skupině na to šel jinak, k nektáriím se prokousával - specialista na jeden druh rostliny). Členové jedné skupiny nikdy pohromadě, ale rozděleni podle gradientu nadmořské výšky. Pokud spolu B. appositus a B.flavifrons, tak každý preferoval jiný druh rostliny. Pokud byl jeden z těchto dvou druhů uměle odstraněn, tak druhý ihned začal využívat „uvolněný“ druh rostliny.
Velikost špičáků u kun - další důkaz rozdělení nik rozrůzněním ústního ústrojí.
Hutchinsonovo empirické „pravidlo“ o poměru velikostí Hutchinson 1959 shromáždil celou řadu příkladů jak od bezobratlých, tak i obratlovců, o velikostních poměrech v škálách potencionálních kompetitorů. Rozdíl tělesné hmotnosti „sousedících“ druhů je roven přibližně dvojnásobku, zatímco délkové míry se lišily relativně 1,3×. Kupodivu to platí i pro lidské výtvory, např. pro běžné dechové nástroje, nebo pro řadu bicyklů od dětských k pro dospělé (Horn & May 1977).
Hutchinsonovo empirické „pravidlo“ o poměru velikostí Kdyby platilo, tak koexistující potencionální kompetitoři budou morfologicky odlišnější, než aby to byla (statistická) náhoda. Jeden z příkladů - Bowers & Brown, 1982 - vztah mezi velikostí těla a společným výskytem hlodavců v pouštních oblastech amerického kontinentu. Uvažovali jen hojné druhy (nad 5% četnosti), ale velké hlodavce nad 80g brali všechny. Pro každou oblast zkoumali 95 lokalit. Hodnocení: pro každou oblast zvlášť a pak pro všechny dohromady spočítali frekvenci výskytu každého druhu. Poté pro každý pár druhů spočítali očekávanou frekvenci společného výskytu (předpoklad - distribuce jsou nezávislé, tedy náhodné). Pro každý pár byl rovněž hodnocen poměr hmotností - hranice konzervativně jako 1.5. Očekávané frekvence pak srovnali se skutečnými. Kdyby platil Hutchinsonův předpoklad, tak druhy stejné velikosti se budou spolu vyskytovat méně často než náhodně, pro druhy s rozdílnou velikostí (poměr nad 1,5) by toto neplatilo.
„Když není kam uhnout“ (když se nika nedá rozdělit či rozšířit) Klasický experiment s kulturami tří druhů trepek (Gause, 1934).
Princip kompetitivního vyloučení Další experimentální důkazy - pokus s potemníky (Park, 1954). V původním pokusu dva faktory - vlhkost a teplota, následovníci přidávali další. Pokud se realizované niky nemohou odlišit, jeden druh potlačí druhého.
Chthalamus Balanus Kompetice dvou druhů vilejšů o prostor Oba obývají přílivovou zónu, ale každý jinde. Přitom Chthalamus, pokud je sám, zabírá mnohem širší pásmo. Balanus je ale „agresivnější“, je schopen rozdrtit usedlé jedince Chthalamus. A ti, co přežijí, nedorůstají takové velikosti jako ti v pásmu bez Balanus. Chthalamus má ale východisko - vydrží být déle na suchu. I mezidruhová kompetice je asymetrická
Kompetice dvou druhů orobince o prostor I mezidruhová kompetice je asymetrická Dva druhy orobince - pokud vysazeny spolu, o.širokolistý preferuje mělčí vodu, o.úzkolistý hlubší (a). Ale pokud rostou každý zvlášť, tak o.úzkolistý má širší niku než o.širokolistý. To se nakonec potvrdilo i po delší době - o.úzkolistý vyhrává.
Kompetice dvou druhů rozsivek o křemík Tilman 1981
Kompetice dvou druhů o zdroj - další experimentální příklady
Jak moc se niky musí lišit, aby umožnily koexistenci? (MacArthur & Levins, 1967, May, 1973) Model - 3 druhy podle gradientu jednoho zdroje. Dva krajní mají stejné K (K1=K3), mezi ně vecpeme další. Jde o to, jak moc vzdálená jsou od sebe optima (d) a jak jsou „nároky“ jednotlivých druhů široké (w). Čím jsou optima blíže (d je malé) a čím jsou nároky širší (w je velké), tím je kompetice vyšší. Poměr d/w a poměr nosných kapacit K1/K2 určuje možnosti koexistence. Model je sice dost imaginární, ale dává konzistentní představu.
Kompetice - modulární organismy (rostliny) Často kontinuální růst - možnost individuální adaptace. Příklad - kompetice stromů o světlo. Po probírce lesa vyšší přírůstky dřevní hmoty.
Co ještě umožňuje koexistenci soutěžících druhů? Variabilita v dostupnosti zdroje Příklad - několik druhů řas v chemostatu (Sommer 1985). P i Si v limitujících koncentracích. Pokud kontinuální přísun P, tak přežívají 1 - 2 druhy. Pokud P dodáván v týdenních pulsech, přežívá druhů výrazně více. První 4 druhy mají schopnost akumulovat P do zásoby. Biomasa dalších dvou druhů kopíruje koncentraci P. Rozsivky ovšem nevydrží - pro ně je limitující Si (respektive poměr Si : P)
Co ještě umožňuje koexistenci soutěžících druhů? Správné načasování Pokud je vstup druhého, byť jinak silnějšího soupeře, opožděn, slabší může získat rozhodující výhodu. „Kdo dřív přijde ten dřív mele“. aneb též Mopolizační hypotéza
Co ještě umožňuje koexistenci soutěžících druhů? Správné načasování 2 Doby kvetení 10 květin z kostarického deštného pralesa - všechny jsou opylovány stejným druhem kolibříka. Aby se o něj nemusely dělit (soutěžit), kvete každá rostlina v jiném období.
Co ještě umožňuje koexistenci soutěžících druhů? Je-li slabší kompetitor rychlejším kolonizátorem
Co ještě umožňuje koexistenci soutěžících druhů? Heterogenita zdroje - zdroj se nevyskytuje rovnoměrně, ale agregovaně - ve shlucích. k je míra agregace, < 5 je agregovaná distribuce Počítačová simulace - koexistence dvou druhů, aniž by došlo k diferenciaci nik. V homogenním prostředí (prázdné symboly) dojde ke kompetitivnímu vyloučení. Pokud má ale zdroj (a tím pádem i kompetitoři) agregovanou distribuci, oba přežívají - byť hůře, než kdyby byli sami. (Atkinson a Shorrocks, 1981).
Výsledek kompetice záleží na evoluční zkušenosti Pokus s jetelem (T) a jílkem(L) Dvě plochy - jedna s oběma druhy, druhá pouze s jetelem. Malé plošky uvnitř - experimentální odstranění jetele (-T) a/nebo jílku (-L). Výsledek v suché váze jetele na plochu (b). V případě (-T: 1,4) replantovaný T roste lépe než transplantovaný (replantovaný má posunutější niku). To potvrzuje případ 5, kdy replantovaný T neroste lépe než když s L (4). Ovšem transplantovaný T (2) není od L diferencovaný, a proto roste daleko lépe. Na ploše se samotným T je to obdobné - původní T roste daleko lépe než T přenesený z „dvoudruhové“ plochy. (Turkington & Mehrhoff, 1990).
Zdánlivá kompetice - soutěž o prostor bez predátora Různé typy kompetičních vztahů - podle výsledku se nedají rozlišit. a) a zejména b) je kompetice „bez přívlastků“ c) a d) řadíme pod tzv. zdánlivou kompetici. Predátor (E) se živí jak druhem C1, tak druhem C2. Proto roste-li populace predátora konzumací druhu C1, tak zvýšeným predačním tlakem trpí i druh C2. Populace druhu C1 tak nepřímo ovlivňuje populaci C2, a samozřejmě to platí i vice versa. Chybí tu tedy limitující zdroj - pokud za něj nepovažujeme prostor bez predátora. C2 a C3 jsou mutualisti
Zdánlivá kompetice - soutěž o prostor bez predátora Nejlepší řešení: odlišit se, aby predátor nebyl společný (= diferenciace nik). A to se dá buď osídlením jiného prostředí, nebo změnou chování. Odlišení nik tedy opět umožňuje koexistenci. Příklady jsou vzácné. Jedním z mála je terénní pokus Schmitta, 1987. Sledoval dvě skupiny kořisti na útesech v přílivové zóně na jednom z kalifornských ostrovů. Kořist: První skupina - pohybliví plži (Tegula a Astraea) Druhá skupina - přisedlí mlži (Chama) Predátoři: Langusta, chobotnice a dravý plž. I když žrali všechny výše zmíněné, tak preferovali mlže. Výchozí stav: Na útesech se členitým reliéfem hodně mlžů, a také hodně predátorů, ale méně plžů . Na „hladkých“ útesech nebyli naopak žádní mlži, ale hodně plžů. Predátorů poskrovnu.
Zdánlivá kompetice - soutěž o prostor bez predátora Plné sloupce = přidané škeble k plžím (hladkým) útesům Prázdné sloupce = kontrola Obě skupiny kořisti se vyskytují recipročně - tam, kde jsou mlži, chybí plži, a naopak. Ale nic v jejich potravě neukazuje na přímou kompetici. Pokud ovšem byli mlži uměle vysazeni na „hladké“ útesy, zvýšil se tam i počet predátorů, a jeho následkem i mortalita plžů - jejich počet poklesl. Plné sloupce = vysoká denzita plžů Prázdné sloupce = nízká denzita plžů Umělé vysazování plžů na členité útesy se kvůli jejich mobilitě nedařilo, ale přesto se dalo pozorovat, že na „hladkých“ útesech s vysokou denzitou plžů bylo více predátorů (bez ohledu na počty mlžů). Na členitých útesech bez mlžů (což byl ovšem vzácný stav) bylo predátorů méně a plžů více než na členitých útesech obývaných mlžem Chama.
Model mezidruhové kompetice 1) Lotka-Volterra Výchozí je logistická rovnice , ale přidá se působení druhého druhu jako ekvivalent vnitrodruhové kompetice. Tvorba ZNGI (Zero Net Growth Isoclines) pro druh 1: r1N1(K1 - N1 - 12N2) = 0 r1 = 0 N1 = 0 K1 - N1 - 12N2 = 0 pro (a) N1 = K1 - 12N2 N1 = 0, N2 = K1 /12 bod A N2 = 0, N1 = K1 bod B pro druh 2 (b) analogicky
Když 12 1 = Když 12 0 = Lotka-Volterra • Ale: • nejjednodušší z modelů, ale biologicky nerealistický • efekt kompetice je lineární • jak měřit v reálných situacích? • Nedefinuje mechanismus kompetice, ale jen její výsledek. INTERAKTIVNÍ MODEL LOTKA-VOLTERRA
Lotka-Volterra Model funguje i pro reálná data, ovšem musí se upravit.
Model mezidruhové kompetice 2) Tilmanův model Mechanistický model. Zdroje mohou být zastupitelné nebo nezastupitelné. V tom rozdíl mezi autotrofy a heterotrofy: U živočichů je potrava zdrojem jak stavebních látek, tak energie (a je většinou zastupitelná), u autotrofů se zdroje vyskytují odděleně, a jsou nezastupitelné. Neastupitelné - esenciální: bez interakce (Liebigův zákon minima) interaktivní Zastupitelné: izoklina protíná osu, protože růst je možný, i když je jeden zdroj nulový. Zcela - kombinace zdrojů nemá žádný +/- efekt. Doplňující se - jejich kombinace je lepší než jen jeden z nich. Antagonistické - jejich kombinace je horší než jen jeden z nich.
A vyhraje – stačí mu méně než druhu B B vyhrává – stačí mu méně než druhu A Tilmanůvmodel Modelový chemostat - jeden zdroj R* - koncentrace zdroje, při které rychlost růstu = 0, velikost populace se nemění.
Modelový chemostat - dva zdroje (nezastupitelné a zastupitelné). Zdroje dodávány v konstantním poměru (, supply point),ale jejich aktuální koncentrace () se může měnit v závislosti na biologických dějích v chemostatu. Pokud je nic nežere, tak to vždy směřuje do supply point - vektor s. Pokud jsou v chemostatu organismy, tak žerou, a to v optimálním poměru (jež závisí na stechio-metrických poměrech prvků v tkáních atd.). Výsledkem je vektor konzumace c. Aktuální koncentrace zdrojů se potom mění ve směru výsledného vektoru vzniklého složením vektorů (s a c). Vektor c má konstantní směr, směr vektoru s je proměnlivý, výsledný vektor je tedy rovněž proměnlivý. Tilmanův model
Tilmanův model • Dva zdroje a dva druhy • 1) záleží, kde je supply point. Pokud je mezi ZNGI a osami, tak žádný druh nepřežije • 2) Pokud jsou ZNGI obou druhů paralelní, vyhrává ten, jehož ZNGI je blíž osám. Pokud se kříží, tak vymezují 6 oblastí: • nikdo nepřežije • přežije druh 1 • přežije druh 2 • oba mohou růst,ale každý žere podle svého vektoru konzumace (c1a c2). Jakmile se aktuální koncentrace přiblíží ZNGI jednoho z druhů, tak ten přestává růst (a tím pádem užírat). Bod aktuální koncentrace se proto začne přesouvat směrem k ZNGI druhého druhu. A díky kombinaci s vektorem s to skončí v průsečíku obou ZNGI.
Tilmanův model To platí, pokud supply point leží mezi c1a c2. Jestliže leží nad c1, tak trajektorie nakonec protne ZNGI druhu 2 a ten je v háji. A vice versa v případě, že supply point leží pod c2. (viz též předchozí barevné schema) Dá se to kombinovat pro více druhů, a jiné kombinace zdrojů. Hranice přežití a vymizení jsou tedy určovány poměrem limitujích zdrojů, a ne jejich absolutní koncentrací. Maximální počet koexistujících druhů je dán počtem limitu-jících zdrojů. Při daném poměru dvou zdrojů mohou přežít vždy jen dva druhy, více druhů jen v případě, že existuje gradient konzumačních poměrů dvou zdrojů (obr. B a D) - průsečík dvou koexistujících musí být v oblasti, kde žádný jiný nemůže přežít. Shrnuto: 1) Koexistující druhy jsou si nejpodobnější (vzhledem k vektoru c), a 2) výsledek kompetice není určen výchozí abundancí.
Tilmanův model - příklady Limit množství potravy (řas) nutný k růstu perlooček r.Daphnia se liší - a ve smyslu Tilmanova modelu zhruba odpovídá „kompetiční síle“ jednotlivých druhů.
Tilmanův model - příklady - vířníci Brachionus rubens a Brachionus calyciflorus
Tilmanův model - příklady Zdroj (teplota) je variabilní Další příklad