Download
1 / 1
10 likes | 155 Views
640.7. OSCILLATIONS OF LOCAL FIELD POTENTIALS IN THE RAT DORSAL HIPPOCAMPUS, DORSAL STRIATUM, AND NUCLEUS ACCUMBENS: COMPARING PLACE AND RESPONSE TRIALS.

E N D
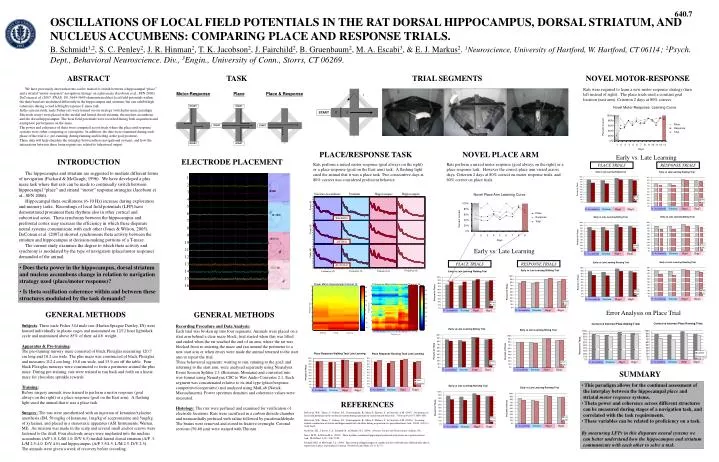
640.7 OSCILLATIONS OF LOCAL FIELD POTENTIALS IN THE RAT DORSAL HIPPOCAMPUS, DORSAL STRIATUM, AND NUCLEUS ACCUMBENS: COMPARING PLACE AND RESPONSE TRIALS. B. Schmidt1,2, S. C. Penley2, J. R. Hinman2, T. K. Jacobson2, J. Fairchild2, B. Gruenbaum2, M. A. Escabi3, & E. J. Markus2.1Neuroscience, University of Hartford, W. Hartford, CT 06114; 2Psych. Dept., Behavioral Neuroscience. Div., 3Engin., University of Conn., Storrs, CT 06269. ABSTRACT TASK TRIAL SEGMENTS NOVEL MOTOR-RESPONSE We have previously shown that rats can be trained to switch between a hippocampal “place” and a striatal “motor-response” navigation strategy on a plus maze (Jacobson et al., SFN 2006). DeCoteau et al. (2007, PNAS, 101:5644-5649) demonstrated that local field potentials within the theta band are modulated differently in the hippocampus and striatum, but can exhibit high coherence during a cued left/right response T-maze task.In the current study, male Fisher rats were trained on our strategy switch plus maze paradigm. Electrode arrays were placed in the medial and lateral dorsal-striatum, the nucleus accumbens, and the dorsal hippocampus. The local field potentials were recorded during both acquisition and asymptotic performance on the maze.The power and coherence of theta were compared across trials where the place and response systems were either competing or synergistic. In addition, the data were examined during each phase of the trial (i.e. pre-running, during running and feeding at the goal position).These data will help elucidate the interplay between these navigational systems, and how the interactions between these brain regions are related to behavioral output. Rats were required to learn a new motor response strategy (turn left instead of right). The place trials used a constant goal location (east arm). Criterion 2 days at 80% correct. Motor-Response Place Place & Response 1 START START Novel Motor-Response Learning Curve 2 3 START 100% 80% Place START START 60% Percent Correct Response 1 2 3 WALKING to start arm SITTING before start RUNNING to goal 40% Total 20% 3 0% START START 1 2 3 4 5 6 7 8 9 10 11 12 13 Days PLACE/RESPONSE TASK NOVEL PLACE ARM Early vs. Late Learning INTRODUCTION ELECTRODE PLACEMENT Rats perform a mixed motor response (goal always on the right) or a place response (goal on the East arm) task. A flashing light cued the animal that it was a place task Two consecutive days at 80% correct was considered proficient behavior. Rats perform a mixed motor response (goal always on the right) or a place response task. However the correct place arm varied across days. Criterion 2 days at 80% correct on motor response trials, and 60% correct on place trials. PLACE TRIALS RESPONSE TRIALS The hippocampus and striatum are suggested to mediate different forms of navigation (Packard & McGaugh, 1996). We have developed a plus maze task where that rats can be made to continually switch between hippocampal “place” and striatal “motor” response strategies (Jacobson et al., SFN 2006). Hippocampal theta oscillations (6-10 Hz) increase during exploration and memory tasks. Recordings of local field potentials (LFP) have demonstrated prominent theta rhythms also in other cortical and subcortical areas. Theta synchrony between the hippocampus and prefrontal cortex may increase the efficiency in which these disparate neural systems communicate with each other (Jones & Wilson, 2005). DeCoteau et al. (2007a) showed synchronous theta activity between the striatum and hippocampus at decision-making portions of a T-maze. The current study examines the degree to which theta activity and synchrony is modulated by the type of navigation (place/motor response) demanded of the animal. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Nucleus Accumbens Striatum Hippocampus Hippocampus Novel Place Arm Learning Curve 50 45 40 100% 35 Power dB 30 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 80% 25 20 Place 15 60% 10 WALKING Response Percent Correct 5 40% Total 50 45 20% 40 35 Power dB 30 0% 1 25 1 2 3 4 5 6 7 8 9 20 15 Days 10 SITTING 5 50 45 Early vs. Late Learning 40 35 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 Power dB 30 25 20 15 PLACE TRIALS RESPONSE TRIALS 10 RUNNING • Does theta power in the hippocampus, dorsal striatum and nucleus accumbens change in relation to navigation strategy used (place/motor response)? • Is theta oscillation coherence within and between these structures modulated by the task demands? 5 0 0 5 10 15 20 25 30 35 40 45 50 5 10 15 20 25 30 35 40 45 50 5 10 15 20 25 30 35 40 45 50 5 10 15 20 25 30 35 40 45 50 Frequency Hz Frequency Hz Frequency Hz Frequency Hz Power Matrix Hippocampal Channel 12 Coherence Matrix Hippocampal Channel 16 50 50 30 0.9 45 45 25 40 0.8 40 20 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 35 0.7 35 15 30 0.6 30 10 Frequency Hz 25 Frequency (Hz) 0.5 25 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 Error Analysis on Place Trial 5 GENERAL METHODS GENERAL METHODS 20 0.4 20 0 15 0.3 15 -5 10 0.2 10 -10 Subjects: Three male Fisher 344 male rats (Harlan Sprague Dawley, IN) were housed individually in plastic cages and maintained on 12/12 hour light/dark cycle and maintained above 85% of their ad lib. weight. Apparatus & Pre-training: The pre-training runway maze consisted of black Plexiglas measuring 120.7 cm long and 10.2 cm wide. The plus maze was constructed of black Plexiglas and measures 112.4 cm long, 10.8 cm wide, and 15.9 cm off the table. Four black Plexiglas runways were constructed to form a perimeter around the plus maze. During pre-training, rats were trained to run back and forth on a linear maze for chocolate sprinkle rewards. Training: Before surgery animals were trained to perform a motor response (goal always on the right) or a place response (goal on the East arm). A flashing light cued the animal that it was a place task. Surgery: The rats were anesthetized with an injection of ketamine/xylazine anesthesia (IM, 50 mg/kg of ketamine, 1mg/kg of acepromazine and 5mg/kg of xylazine), and placed in a stereotaxic apparatus (ASI Instruments, Warren, MI). An incision was made in the scalp and several small anchor screws were fastened to the skull. Four electrode arrays were implanted into the nucleus accumbens (A/P 1.0, L/M 1.0, D/V 6.5) medial-lateral dorsal striatum (A/P .3, L/M 2.5-4.0, D/V 4.0) and hippocampus (A/P 3.5/4.5, L/M 2.5, D/V 2.5). The animals were given a week of recovery before recording. Recording Procedure and Data Analysis: Each trial was broken up into four segments: Animals were placed on a start arm behind a clear maze block; trial started when this was lifted and ended when the rat reached the end of an arm; where the rat was blocked from re-entering the maze and ran around the perimeter to a new start arm or when errors were made the animal returned to the start arm to repeat the trial. Three behavioral segments: waiting to run; running to the goal; and returning to the start arm, were analysed separately using Neuralynx Event Session Splitter 2.1 (Bozeman, Montana) and converted into wav format using Neuralynx CSC to Wav Audio Converter 2.1. Each segment was concatenated relative to its trial type (place/response, competitive/cooperative) and analyzed using MatLab (Natick, Massachusetts). Power spectrum densities and coherence values were measured. Histology: The rats were perfused and examined for verification of electrode locations. Rats were sacrificed in a carbon dioxide chamber and transcardially perfused with saline followed by parafomaldehyde. The brains were removed and stored in fixative overnight. Coronal sections (50-60 m) were stained with Thionin. 5 0.1 5 -15 0 0 1 2 3 4 5 6 7 8 9 10 11 13 14 15 Walking Sitting Running Channel (x,y) N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 SUMMARY • This paradigm allows for the continual assessment of the interplay between the hippocampal place and striatal motor response systems. • Theta power and coherence across different structures can be measured during stages of a navigation task, and correlated with the task requirements. • These variables can be related to proficiency on a task. • By measuring LFPs in this disparate neural systems we can better understand how the hippocampus and striatum communicate with each other to solve a task. N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2 REFERENCES DeCoteau, W.E., Thorn, C., Gibson, D.J., Courtemanche, R., Mitra, P., Kubota, Y., & Graybiel, A.M. (2007). Oscillations of local field potentials in the rat dorsal striatum during spontaneous and instructed behaviors. J Neurophysiol 97:3800-3805. DeCoteau, W.E., Thorn, C., Gibson, D.J., Courtemanche, R., Mitra, P., Kubota, Y., & Graybiel, A.M. (2007a). Learning-related coordination of striatal and hippocampal theta rhythms during acquisition of a procedural maze task. PNAS, 104(13), 5644-5649. Jacobson, T.K., Cleaver, C.A., Schmidt, B., & Markus, E.J. (2006). Abstract. Society for Neuroscience. Atlanta, GA. Jones, M.W., & Wilson M.A. (2005). Theta rhythms coordinate hippocampal-prefrontal interactions in a spatial memory task. PLoS Biol, 3(12), 2187-2199. Packard, M.G. & McGaugh, J.L. (1996). Inactivation of hippocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning. Neurbiol Learn Mem, 65(1): 65-72. N. Accumbens Striatum Hipp1 Hipp2 N. Accumbens Striatum Hipp1 Hipp2