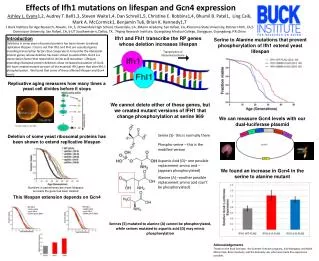
Download

1 / 55
550 likes | 675 Views
Limits on Human Lifespan and Molecular Effects on Ageing. Leonid A. Gavrilov Natalia S. Gavrilova Center on Aging, NORC/University of Chicago, 1155 East 60th Street, Chicago, IL 60637. Questions of Scientific and Practical (Actuarial) Significance. How far could mortality decline go?

E N D
Limits on Human Lifespan and Molecular Effects on Ageing Leonid A. Gavrilov Natalia S. Gavrilova Center on Aging, NORC/University of Chicago, 1155 East 60th Street, Chicago, IL 60637
Questions of Scientific and Practical (Actuarial) Significance • How far could mortality decline go? (absolute zero seems implausible) • Are there any ‘biological’ limits to human mortality decline, determined by ‘reliability’ of human body? (lower limits of mortality dependent on age, sex, and population genetics) • Were there any indications for ‘biological’ mortality limits in the past? • Are there any indications for mortality limits now?
The Gompertz-Makeham Law μ(x) = A + R0exp(α x) A – Makeham term or background mortality R0exp(α x) – age-dependent mortality
Historical Changes in Mortality for 40-year-old Swedish Males • Total mortality • Background mortality • Age-dependent mortality • Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Historical Changes in Mortality for 40-year-old Women in Norway and Denmark • Norway, total mortality • Denmark, total mortality • Norway, age-dependent mortality • Denmark, age-dependent mortality Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Historical Changes in Mortality for 40-year-old Italian Women and Men • Women, total mortality • Men, total mortality • Women, age-dependent mortality • Men, age-dependent mortality Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Historical Changes in Survival from Age 90 to 100 years. France
Historical Changes in Survival from Age 90 to 100 years. Japan
Extension of the Gompertz-Makeham Model through the Factor Analysis of Mortality Trends Mortality force (age, time) = = a0(age) + a1(age) x F1(time) + a2(age) x F2(time)
Preliminary Conclusions • There was some evidence for ‘ biological’ mortality limits in the past, but these ‘limits’ proved to be responsive to the recent technological and medical progress. • Thus, there is no convincing evidence for absolute ‘biological’ mortality limits now. • Analogy for illustration and clarification:There was a limit to the speed of airplane flight in the past (‘sound’ barrier), but it was overcome by further technological progress. Similar observations seems to be applicable to current human mortality decline.
Molecular Effects on Ageing New Ideas and Findings by Bruce Ames: • The rate of mutation damage is NOT immutable, but it can be dramatically decreased by very simple measures: -- Through elimination of deficiencies in vitamins and other micronutrients (iron, zinc, magnesium, etc). • Micronutrient deficiencies are very common even in the modern wealthy populations • These deficiencies are much more important than radiation, industrial pollution and most other hazards Our hypothesis: Remarkable improvement in the oldest-old survival may reflect an unintended retardation of the aging process, caused by decreased damage accumulation, because of improving the micronutrient status in recent decades
Micronutrient Undernutrition in Americans Nutrient Population Group % ingesting <RDA <50% RDA % ingesting < 50% RDA RDA Minerals Iron Women20-30 years 18 mg 75% 25% Women 50+ years 8 mg 25% 5-10% Men; Women 50+ years 11; 8 mg 10% 50% Zinc Vitamins B6 Men; Women 1.7; 1.5 mg 50% 10% 75% Folate** Men; Women 400 mcg 25%; 50% B12 Men; Women 2.4 mcg 10-20; 25-50 % 5; ~10-25% C Men; Women 90; 75 mg 50% 25% •Wakimoto and Block (2001) J Gerontol A Biol Sci Med Sci. Oct; 56 Spec No 2(2):65-80. ** Before U.S. Food FortificationSource: Presentation by Bruce Ames at the IABG Congress
Molecular Effects on Ageing (2) Ideas and Findings by Bruce Ames: • The rate of damage accumulation is NOT immutable, but it can be dramatically decreased by PREVENTING INFLAMMATION: Inflammation causes tissue damage through many mechanisms including production of Hypochlorous acid (HOCl), which produces DNA damage (through incorporation of chlorinated nucleosides). Chronic inflammation may contribute to many age-related degenerative diseases including cancer Hypothesis: Remarkable improvement in the oldest-old survival may reflect an unintended retardation of the aging process, caused by decreased damage accumulation, because of partial PREVENTION of INFLAMMATION through better control over infectious diseases in recent decades
Characteristic of our Dataset • Over 16,000 persons belonging to the European aristocracy • 1800-1880 extinct birth cohorts • Adult persons aged 30+ • Data extracted from the professional genealogical data sources including Genealogisches Handbook des Adels, Almanac de Gotha, Burke Peerage and Baronetage.
Season of Birth and Female Lifespan8,284 females from European aristocratic families born in 1800-1880Seasonal Differences in Adult Lifespan at Age 30 • Life expectancy of adult women (30+) as a function of month of birth (expressed as a difference from the reference level for those born in February). • The data are point estimates (with standard errors) of the differential intercept coefficients adjusted for other explanatory variables using multivariate regression with categorized nominal variables.
Season of Birth and Female Lifespan6,517 females from European aristocratic families born in 1800-1880Seasonal Differences in Adult Lifespan at Age 60 • Life expectancy of adult women (60+) as a function of month of birth (expressed as a difference from the reference level for those born in February). • The data are point estimates (with standard errors) of the differential intercept coefficients adjusted for other explanatory variables using multivariate regression with categorized nominal variables.
Mean Lifespan of FemalesBorn in December and Februaryas a Function of Birth Year • Life expectancy of adult women (30+) as a function of year of birth
Daughters' Lifespan (30+) as aFunctionof Paternal Age at Daughter's Birth6,032 daughters from European aristocratic familiesborn in 1800-1880 • Life expectancy of adult women (30+) as a function of father's age when these women were born (expressed as a difference from the reference level for those born to fathers of 40-44 years). • The data are point estimates (with standard errors) of the differential intercept coefficients adjusted for other explanatory variables using multiple regression with nominal variables. • Daughters of parents who survived to 50 years.
Daughters' Lifespan (60+) as aFunctionof Paternal Age at Daughter's Birth4,832 daughters from European aristocratic familiesborn in 1800-1880 • Life expectancy of older women (60+) as a function of father's age when these women were born (expressed as a difference from the reference level for those born to fathers of 40-44 years). • The data are point estimates (with standard errors) of the differential intercept coefficients adjusted for other explanatory variables using multiple regression with nominal variables. • Daughters of parents who survived to 50 years.
Paternal Age as a Risk Factor for Alzheimer Disease • MGAD - major gene for Alzheimer Disease • Source: L. Bertram et al. Neurogenetics, 1998, 1: 277-280.
Paternal Age and Risk of Schizophrenia • Estimated cumulative incidence and percentage of offspring estimated to have an onset of schizophrenia by age 34 years, for categories of paternal age. The numbers above the bars show the proportion of offspring who were estimated to have an onset of schizophrenia by 34 years of age. • Source: Malaspina et al., Arch Gen Psychiatry.2001.
What Should the Aging Theory Explain: • Why do most biological species deteriorate with age? • Specifically, why do mortality rates increase exponentially with age in many adult species (Gompertz law)? • Why does the age-related increase in mortality rates vanish at older ages (mortality deceleration)? • How do we explain the so-called compensation law of mortality (Gavrilov & Gavrilova, 1991)?
Exponential Increase of Death Rate with Age in Fruit Flies(Gompertz Law of Mortality) Linear dependence of the logarithm of mortality force on the age of Drosophila. Based on the life table for 2400 females of Drosophila melanogaster published by Hall (1969). Mortality force was calculated for 3-day age intervals. Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Age-Trajectory of Mortality in Flour Beetles(Gompertz-Makeham Law of Mortality) Dependence of the logarithm of mortality force (1) and logarithm of increment of mortality force (2) on the age of flour beetles (Tribolium confusum Duval). Based on the life table for 400 female flour beetles published by Pearl and Miner (1941). Mortality force was calculated for 30-day age intervals. Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Age-Trajectory of Mortality in Italian Women(Gompertz-Makeham Law of Mortality) Dependence of the logarithm of mortality force (1) and logarithm of increment of mortality force (2) on the age of Italian women. Based on the official Italian period life table for 1964-1967. Mortality force was calculated for 1-year age intervals. Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Compensation Law of MortalityConvergence of Mortality Rates with Age 1 – India, 1941-1950, males 2 – Turkey, 1950-1951, males 3 – Kenya, 1969, males 4 - Northern Ireland, 1950-1952, males 5 - England and Wales, 1930-1932, females 6 - Austria, 1959-1961, females 7 - Norway, 1956-1960, females Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Compensation Law of Mortality in Laboratory Drosophila 1 – drosophila of the Old Falmouth, New Falmouth, Sepia and Eagle Point strains (1,000 virgin females) 2 – drosophila of the Canton-S strain (1,200 males) 3 – drosophila of the Canton-S strain (1,200 females) 4 - drosophila of the Canton-S strain (2,400 virgin females) Mortality force was calculated for 6-day age intervals. Source: Gavrilov, Gavrilova, “The Biology of Life Span” 1991
Mortality at Advanced Ages Source:Gavrilov L.A., Gavrilova N.S. The Biology of Life Span: A Quantitative Approach, NY: Harwood Academic Publisher, 1991
Survival Patterns After Age 90 Percent surviving (in log scale) is plotted as a function of age of Swedish women for calendar years 1900, 1980, and 1999 (cross-sectional data). Note that after age 100, the logarithm of survival fraction is decreasing without much further acceleration (aging) in almost a linear fashion. Also note an increasing pace of survival improvement in history: it took less than 20 years (from year 1980 to year 1999) to repeat essentially the same survival improvement that initially took 80 years (from year 1900 to year 1980). Source: cross-sectional (period) life tables at the Berkeley Mortality Database (BMD): http://www.demog.berkeley.edu/~bmd/
Non-Gompertzian Mortality Kinetics of Four Invertebrate Species Non-Gompertzian mortality kinetics of four invertebrate species: nematodes, Campanularia flexuosa, rotifers and shrimp. Source: A. Economos. A non-Gompertzian paradigm for mortality kinetics of metazoan animals and failure kinetics of manufactured products. AGE, 1979, 2: 74-76.
Non-Gompertzian Mortality Kinetics of Three Rodent Species Non-Gompertzian mortality kinetics of three rodent species: guinea pigs, rats and mice. Source: A. Economos. A non-Gompertzian paradigm for mortality kinetics of metazoan animals and failure kinetics of manufactured products. AGE, 1979, 2: 74-76.
Non-Gompertzian Mortality Kinetics of Three Industrial Materials Non-Gompertzian mortality kinetics of three industrial materials: steel, industrial relays and motor heat insulators. Source: A. Economos. A non-Gompertzian paradigm for mortality kinetics of metazoan animals and failure kinetics of manufactured products. AGE, 1979, 2: 74-76.
Redundancy Creates Both Damage Tolerance and Damage Accumulation (Aging)
Differences in reliability structure between (a) technical devices and (b) biological systems
Statement of the HIDL hypothesis:(Idea of High Initial Damage Load ) "Adult organisms already have an exceptionally high load of initial damage, which is comparable with the amount of subsequent aging-related deterioration, accumulated during the rest of the entire adult life." Source: Gavrilov, L.A. & Gavrilova, N.S. 1991. The Biology of Life Span: A Quantitative Approach. Harwood Academic Publisher, New York.
Why should we expect high initial damage load ? • General argument:-- In contrast to technical devices, which are built from pre-tested high-quality components, biological systems are formed by self-assembly without helpful external quality control. • Specific arguments: • Cell cycle checkpoints are disabled in early development (Handyside, Delhanty,1997. Trends Genet. 13, 270-275 ) • extensive copy-errors in DNA, because most cell divisions responsible for DNA copy-errors occur in early-life (loss of telomeres is also particularly high in early-life) • ischemia-reperfusion injury and asphyxia-reventilation injury during traumatic process of 'normal' birth
Spontaneous mutant frequencies with age in heart and small intestine Source: Presentation of Jan Vijg at the IABG Congress, Cambridge, 2003
Birth Process is a Potential Source of High Initial Damage • During birth, the future child is deprived of oxygen by compression of the umbilical cord and suffers severe hypoxia and asphyxia. Then, just after birth, a newborn child is exposed to oxidative stress because of acute reoxygenation while starting to breathe. It is known that acute reoxygenation after hypoxia may produce extensive oxidative damage through the same mechanisms that produce ischemia-reperfusion injury and the related phenomenon, asphyxia-reventilation injury. Asphyxia is a common occurrence in the perinatal period, and asphyxial brain injury is the most common neurologic abnormality in the neonatal period that may manifest in neurologic disorders in later life.
Practical implications from the HIDL hypothesis: "Even a small progress in optimizing the early-developmental processes can potentially result in a remarkable prevention of many diseases in later life, postponement of aging-related morbidity and mortality, and significant extension of healthy lifespan." "Thus, the idea of early-life programming of aging and longevity may have important practical implications for developing early-life interventions promoting health and longevity." Source: Gavrilov, L.A. & Gavrilova, N.S. 1991. The Biology of Life Span: A Quantitative Approach. Harwood Academic Publisher, New York.
Failure Kinetics in Mixtures of Systems with Different Redundancy LevelsInitial Period The dependence of logarithm of mortality force (failure rate) as a function of age in mixtures of parallel redundant systems having Poisson distribution by initial numbers of functional elements (mean number of elements, = 1, 5, 10, 15, and 20.
Daughter's Lifespan(Mean Deviation from Cohort Life Expectancy)as a Function of Paternal Lifespan • Offspring data for adult lifespan (30+ years) are smoothed by 5-year running average. • Extinct birth cohorts (born in 1800-1880) • European aristocratic families. 6,443 cases
Offspring Lifespan at Age 30 as a Function of Paternal LifespanData are adjusted for other predictor variables Daughters, 8,284 cases Sons, 8,322 cases
Offspring Lifespan at Age 60 as a Function of Paternal LifespanData are adjusted for other predictor variables Daughters, 6,517 cases Sons, 5,419 cases
Offspring Lifespan at Age 30 as a Function of Maternal LifespanData are adjusted for other predictor variables Daughters, 8,284 cases Sons, 8,322 cases